Учебник по биологии
Рис. 19
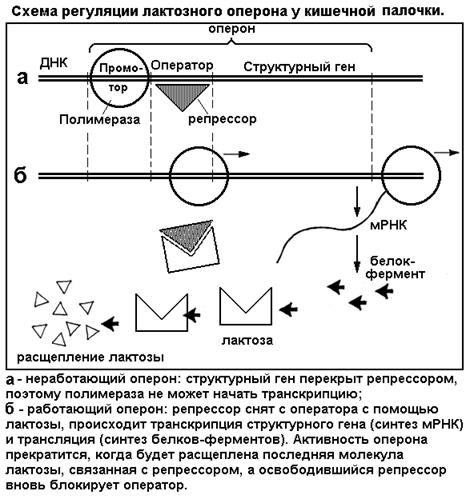
Лактозный оперон - участок молекулы ДНК - состоит из трех частей: промотора, оператора и структурного гена. Промотор - стартовый участок гена, сюда садится фермент РНК-полимераза, ведущий транскрипцию. Оператор - пусковой барьер, в отсутствие лактозы закрытый специальным белком-репрессором. Структурный ген (точнее - здесь находится цепочка, семейство генов) - основной участок ДНК, кодирующий и производящий через иРНК нужный белок-фермент. Пока оператор связан с белком-репрессором, полимераза не может стартовать и структурный ген не работает, синтез фермента отсутствует (см. рис. 19 а). Когда в клетку попадает лактоза, одна ее молекула связывается с репрессором и отнимает его от оператора. Теперь путь полимеразе открыт, идет синтез иРНК (транскрипция) и, далее, синтез соответствующего белка-фермента (трансляция) (рис. 19 б). Ферменты расщепляют поступивший в клетку сахар и в последнюю очередь ту его молекулу, которая связана с репрессором. Но когда будет переварена и эта последняя молекула, белок-репрессор освобождается и вновь блокирует оператор. Производство иРНК и фермента прекращается до поступления новой порции сахара. По своей простоте система регуляции гена концентрацией субстрата похожа на простые технические регуляторы. Напомним, однако, что у эукариот регуляция генной активности более сложная. Она включает возбуждение клеточных рецепторов гормонами или другими биологически активными веществами, запуск каскада реакций вторичных мессенджеров, которые поступают в ядро и избирательно активируют гены.
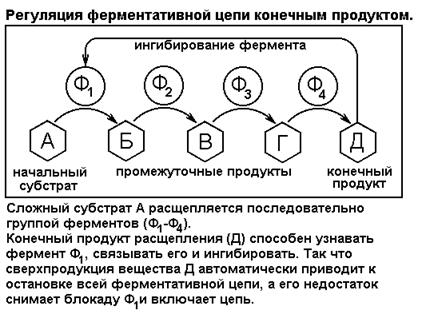
Другой пример простых саморегулирующихся систем, использующих обратную отрицательную связь, представляют ферментативные цепи, ингибируемые конечным продуктом (рис. 20). Такие цепи обычно локализуются на поверхности внутриклеточных или наружных мембран и проводят комплексную переработку сложного субстрата в простой продукт. Суть регуляции состоит в том, что конечный продукт имеет стереохимическое сродство с первым ферментом. Связываясь с ферментом, продукт ингибирует (подавляет) его активность, так как полностью искажает его третичную структуру. Работает следующий регуляторный цикл. При повышении концентрации конечного продукта выше необходимого уровня его избыток ингибирует ферментную цепь (для этого достаточно остановить самый первый фермент). Ферментация прекращается, а свободный продукт расходуется на нужды клетки. Через некоторое время возникает дефицит продукта, блок с ферментов снимается, цепь активируется, и производство продукта снова растет.
Рис. 20
Третий пример- поддержание внутриклеточного осмотического гомеостаза. В сегменте 19 мы говорили о механизме возникновения нервных импульсов и отмечали важную роль ионов натрия, концентрация которых снаружи клетки должна поддерживаться на более высоком уровне, чем внутри. Благодаря натриевым насосам, встроенным в мембрану клетки, удерживается нужный градиент ионов. Как только клетка получает избыток натрия, активируется натриевый насос (его фермент, расщепляющий АТФ и дающий энергию). Натрий выкачивается, его концентрация в клетке падает, что служит сигналом для отключения насоса.
Аналогично в клетках растений с помощью плазмалеммы (наружной мембраны) и вакуолей регулируется состав солей и питательных веществ. Плазмалемма обеспечивает приток в клетку необходимых ионов и воды из внешней среды и выделение балластных и избыточных ионов водорода, натрия, кальция. Мембрана вакуоли регулирует поступление в протоплазму запасных субстратов из вакуоли при их недостатке и удаление в вакуоль - при избытке.
Во всех рассмотренных случаях действует один и тот же принцип - саморегуляция системы на основе обратной отрицательной связи. Заметим, однако, что регулируемые параметры - концентрация солей, питательных веществ, конечного продукта ферментации или продукта генной активности - не бывают абсолютно постоянными, они поддерживаются в допустимых границах. В каждом случае это свои физиологические границы, позволяющие нормально осуществлять клеточные функции. Аналогичный принцип мы увидим и на организменном уровне.
СЕГМЕНТ 28. САМОРЕГУЛЯЦИЯ МНОГОКЛЕТОЧНОГО ОРГАНИЗМА
Как мы только что видели, уже на клеточном уровне возникает необходимость поддержания специфических физико-химических условий, отличающихся от условий окружающей среды. У многоклеточных организмов появляется внутренняя среда, в которой находятся клетки различных органов и тканей, происходит усложнение и совершенствование механизмов гомеостаза. В ходе эволюции формируются специализированные органы кровообращения, дыхания, пищеварения, выделения и др., участвующие в поддержании гомеостаза.
У морских беспозвоночных имеются механизмы стабилизации объема, ионного состава и рН жидкостей внутренней среды. Для животных, перешедших к жизни в пресных водах и на суше, а также у позвоночных, мигрировавших из пресных вод в море, сформированы механизмы осморегуляции, обеспечивающие постоянство концентрации солей внутри организма.
Наиболее совершенен гомеостаз у млекопитающих, что способствует расширению возможностей их приспособления к окружающей среде. В частности, обеспечивается постоянство объема крови и других внеклеточных жидкостей, концентрации в них ионов, осмотически активных веществ, постоянство рН крови, состава в ней белков, липидов и углеводов. У млекопитающих, а также у птиц, в узких пределах регулируется температура тела - их называют теплокровными животными.
Основную роль в поддержании гомеостаза организма играют нервная и гормональная системы регуляции (см. рис. 17 г).
Наиболее важную интегрирующую функцию выполняет центральная нервная система, особенно кора головного мозга. Большое значение имеет и вегетативная нервная система, в частности ее симпатический отдел - система ганглиев (скоплений нервных клеток), расположенных по бокам позвоночника, в брыжейке и других частях тела (например, солнечное сплетение). Чувствительные нервные волокна охватывают сетью все внутренние органы, кровеносные сосуды, обеспечивая рефлекторную взаимосвязь между ними.
Гормональная регуляция обеспечивается системой эндокринных желез (от греческих endon - внутрь и krino - выделяю) - желез внутренней секреции. Центральная эндокринная железа - гипофиз - находится в голове и имеет прямую связь с головным мозгом (через посредство гипоталамуса), а ее гормоны через кровь воздействуют на все местные эндокринные железы- такие как щитовидная, паращитовидная, надпочечники, а также скопления эндокринных клеток в поджелудочной и слюнной железах, семенниках, яичниках, тимусе, плаценте и даже в сердце, желудке, кишечнике, почках. Выделяемые эндокринными железами гормоны с током крови (гуморально) распространяются ко всем органам-мишеням и участвуют в регуляции их роста (см. сегмент 25) и функционирования. Таким образом, фактически благодаря связи нервной и эндокринной систем осуществляется единая нейрогормональная саморегуляция организма.