Популяционно-генетическая структура вида у гидробионтов
Ne, лучше объясняет фактические данные (Рис. 5). Полиморфизм по LDH* является селективно нейтральным и зависит только от двух параметров - Ne и m (Рис. 5).
Классификация вариантов внутривидовой структуры, на основе использования популяционно-генетических средств.
Если основываться на средствах популяционной генетики, то можно определить три главных варианта популяционной организации: (1) панмиктическая популяция, (2) подразделенная популяция и (3) непрерывная популяция. Третий вариант достаточно редок в природе.
(1) Панмиктическая популяция. Этот вариант популяционной организации теоретически описывается моделью Харди-Вайнберга. Внутри такой популяции не существует генетической дифференциации. Поэтому, когда нет дифференцирующего действия отбора, то выполняется условие Fst = 0. Вид может быть представлен на ареале одной или несколькими популяциями, которые полностью, или почти полностью, изолированы одна от другой.
(2) Подразделенная популяция. Теоретически описана как островная модель популяционной структуры (Wright, 1943), или как модель ступенчатой миграции (Kimura, Ohta, 1971). Дифференциация в подразделенной популяции в равновесном состоянии, в отсутствие отбора, зависит, в основном, от двух факторов – миграции и эффективной численности (размера) приближенно через соотношение: Fst = 1 / [4Nm + 1].
Особенности популяционной организации морских организмов.
Имеются некоторые особенности внутривидовой структурированности в морской среде. Это связано с обычно весьма многочисленными популяциями, которые имеют огромный потенциал миграции или расселения (активного или пассивного). Однако все генетические принципы также сохраняют свою верность и для этих популяций. Например, имеются в среднем большие и малые значения Fst для разных видов, что связано, главным образом, с большой и небольшой генетической дифференциацией, зависимой от обратно пропорционально связанных дрейфа и миграции.
Генетическая изменчивость и дифференциация.
Средняя гетерозиготность на особь, H широко варьирует в таксонах растений и животных. Суммарная средняя составила: H=0.076, для позвоночных - H=0.054 и для беспозвоночных - H=0.100 (Nevo et al., 1984).
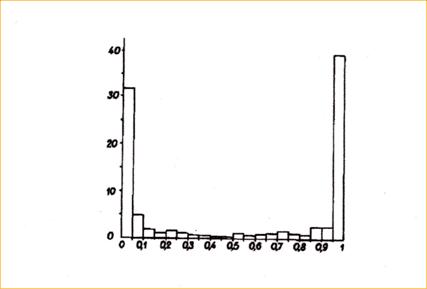
Частоты коэффициентов генетического сходства по ферментным локусам, оцененные для разных видов, следуют U-образному распределению; последнее обратно связано с ожидаемой дифференциацией при нейтральности (Ayala, 1975; рис. 6).
| |||
|
Рис. 6. Ось Y – частота (%), ось X – коэффициент сходства, I среди видов группы Drosophila willistoni. Большинство локусов обнаруживают либо фиксированное, либо нулевое сходство, тогда как остальные варьируют в диапазоне от малых до больших коэффициентов сходства. U-образное распределение не согласуется с дрейфовой дифференциацией при нейтральности белковых маркеров. Распределение должно быть близким к нормальному (По Ayala, 1975). |
Тем не менее, для некоторых белковых локусов можно обнаружить распределение коэффициентов сходства, которое близко к нормальному (Рис. 7). Таким образом, по меньшей мере, по некоторым локусам обнаруживаемая дифференциация во времени согласуется с дрейфовым, нейтральным процессом.
|
|
|
Рис. 7. Частотное распределение коэффициента сходства, рассчитанного по дуплицированным гемоглобиновым локусам лососевых рыб (По Картавцев, 1995). |

Некоторые исследования показывают, что без привлечения концепции естественного отбора вряд ли возможно объяснить сопряженную изменчивость индивидуальной гетерозиготности по ферментным генам (Ho) и морфологической изменчивости в популяционно-средовом градиенте (Рис. 8).
|
|
Рис. 8. График средних, который иллюстрирует результаты двухфакторного дисперсионного анализа: Ho и изменчивости морфологических признаков у брюхоногого моллюска (Nucella heyseana) в заливе Восток и заливе Находка (залив Петра Великого, Японское море). Увеличение размера тройных гетерозигот (Ho =3) в двух выборках трудно объяснить без привлечения концепции отбора(По Kartavtsev, Svinyna, 2003). |

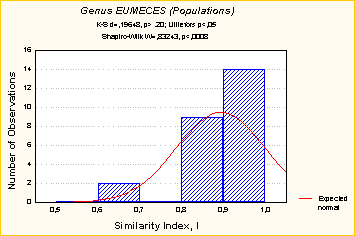
Как продемонстрировано ранее, источники, поддерживающие генетическую изменчивость в популяциях, имеют комплексную природу. Иногда совершенно очевидно, что ни естественный отбор, ни нейтральность не являются причиной того или иного типа обнаруженной изменчивости. Неоднородность самого материала и соответствующих данных могут вести к корреляциям или атипичным распределениям (Рис. 9).
|
|
Рис 9. Частотное распределение коэффициента сходства (I) внутри видов. Верхний график показывает распределение I для всей базы данных, тогда как нижний только для рептилий рода Eumeces (Картавцев, не опубликовано). |
|
|
Очевидна ненормальность обоих распределений. Бимодальность на нижнем графике обусловлена гетерогенностью данных, представленных небольшим числом очень несходных островных популяций этих ящериц (I = 0.6-0.7 и менее; Kato et al., 1994) (Рис. 8.3.7). Эти данные предполагают, что имеется гетерогенность среди популяционных групп по степени сходства и они дифференциально представлены в материале по внутривидовой изменчивости. |
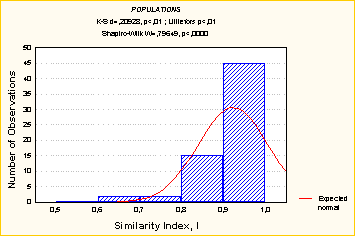
Если сравнивать более однородные группы, то нормальность можно обнаружить (Рис.10).
|
|
Рис. 10. Частотное распределение коэффициента сходства (Rc) внутри вида тихоокеанская сельдь Clupea pallasi (Перерасчет по данным Картавцев, Рыбникова, 1999). |
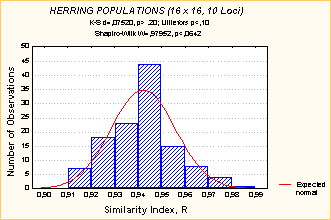
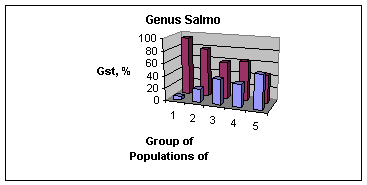
Данные о внутривидовой дифференциации показывают, что единообразия популяционной дифференциации не наблюдается ни у форелей рода Salmo, ни у тихоокеанских лососей рода Oncorhynchus. Хотя можно сделать вывод, что в обеих группах преобладает внутривидовой компонент (Рис. 11).
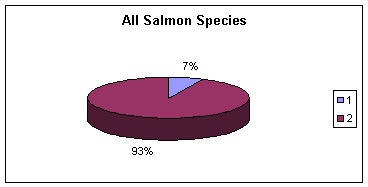 |
Рис. 11. Величина Gst (Nei, 1973, 1977b) как мера внутривидовой дифференциации лососевых рыб (Данные автора, не опубликованы). Данные на Рис. 8.3.9 иллюстрируют: (1) относительно небольшую межпопуляционную генетическую дифференциацию (Gst = 7%, верхний рисунок)) по сравнению с внутрипопуля-ционным разнообразием | ||
|
|
(Gst = 93%) (верхний рисунок) и (2) различие величин Gst между родами Salmo и Oncorchynchus, с одной стороны, и между видами в каждом таксоне (средний и нижний рисунки). Виды и популяции у форелей более дифференцированы генетически, чем у тихоокеанских лососей, о чем свидетельствуют статистически | ||
|
|
значимые различия Gst. Причины, обусловливающие различные уровни генетической дифференциации, могут быть различными. Как отмечалось, одной из самых важных причин может быть различие эффективной численности популяций этих двух групп. |