Липиды центральной нервной системы и структура клеточных мембран
В области температурных фазовых переходов таких липидов отмечается изменение каталитических и транспортных свойств белков. Общая доля кольцевых липидов довольно велика – около 20%. Доказано, что можно изменять активность мембранных белков изменением связанных с ними липидов.
Другой пул липидов, удаленных от белков и подвергающихся быстрой латеральной диффузии, характерной для билипидного слоя, не пронизанного белком, составляет около 80%. Действие этих липидов на мембранные белки аналогично растворяющему эффекту воды на свойства растворимого белка.
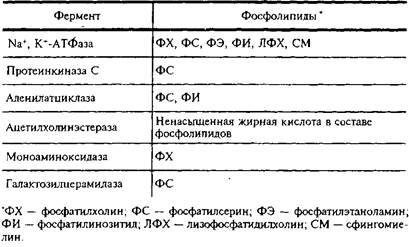
Приведем примеры функциональной роли индивидуальных липидов в мембранах ЦНС. В табл. 5 представлены данные о влиянии различных фосфолипидов на активность мембранных ферментов.
Активирование отдельными фосфолипидами мембранных ферментов
6. УЧАСТИЕ ЛИПИДОВ В РЕЦЕПЦИИ И ПЕРЕДАЧЕ ВНУТРЬ КЛЕТКИ СИГНАЛА
Межклеточные контакты, без которых немыслима деятельность ЦНС, обеспечивают постоянную передачу информации через плазматическую мембрану. Эта передача не может не касаться билипидного слоя. Процесс передачи информации через мембрану включает рецепцию внешнего химического сигнала и трансформацию его во внутриклеточный эффект.
Возникает вопрос, принимают ли участие липиды бислоя в рецепции внешних сигналов. В последнее десятилетие установлено, что сульфоиереброзиды играют довольно специфическую роль в рецепции опиоидов. Частично очищенный препарат рецептора опиоидов содержит высокую концентрацию сульфатидов. Предполагают, что сульфатная группа це-реброзидсульфата входит в состав или соседствует с активным центром опиатных рецепторов, который имеет белковую природу. Возможно, что взаимодействие опиатов с цереброзидсульфатами выполняет вспомогательную функцию, способствуя сосредоточению лигандов в области центра белковой природы.
При исследовании ряда кислых липидов только сульфатиды проявляли наивысшее сродство к опиатам в различных физиологических условиях. Доказательством важной роли сульфати-дов в рецепции опиоидов может служить и тот факт, что антитела к цереброзидсульфату, введенные в мозг крысы, снимали наркотическое действие морфина.
Если липиды бислоя могут быть участниками процесса рецепции, то естественно ожидать их участия в каскаде реакций, возникающих после активации рецепторов. М.Н. Хокин и Л. Э. Хокин впервые связали холинергическую стимуляцию с усилением обмена фосфатидилинозита и фосфатидной кислоты. Явление получило название «фосфолипидного эффекта»; этот термин сейчас заменен на термин «фосфоинозитид-ный эффект», поскольку появилось большое число работ, показывающих именно их регуляторную роль в транспорте вторичного мессенджера – ионов кальция – через мембраны.
Содержание фосфоинозитидов в мембранах ЦНС не превышает 0,5–2% от общих липидов, локализованы они преимущественно в плазматических мембранах, в миелине, в эндоплазматическом ретикулуме, наружной митохондриальной и ядерной мембранах. В состав фосфоинозитидов входит арахидоновая кислота, являющаяся важным источником простагландинов. Деполяризация мембраны приводит к быстрому высвобождению арахидо-новой кислоты именно из фосфоинозитидов. Р. Митчелл высказал гипотезу о прямой связи расщепления фосфоинозитидов с рецепторным аппаратом клетки и увеличением внутриклеточной активности. В синаптосомах стимуляция части мускариновых и ах-адренерги-ческих рецепторов обусловливает фосфоинозитидный эффект, сопровождающийся изменением проницаемости плазматической мембраны для ионов кальция.
Участие фосфоинозитидов и продуктов их обмена в регуляции транспорта кальция осуществляется несколькими путями:
1) при распаде фосфатидилинозитидов образуется 1,2 – диа-цилглицерин, стимулирующий активность протеинкиназы С, которая, в свою очередь, фосфорилирует белок Са-каналов и некоторые другие белки;
2) трифосфоинозитол, освобождающийся при расщеплении фосфатидилинозитидов, обладает высокой способностью связывать двухвалентные катионы; по этой причине он индуцирует мобилизацию мембранно-связанного кальция;
3) инозитол-трифосфат способен также повышать уровень внутриклеточного кальция за счет открытия кальциевых каналов эндоплазматического ретикулума. Таким образом, происходит сопряжение выброса кальция из внутриклеточных мест хранения с входом кальция через мембраны.
До включения описанного механизма концентрация свободного кальция в цитоплазме нейрона составляет примерно 1 – .10~М. Концентрация кальция снаружи нейрона в десятки тысяч раз выше. Мобилизация Са+ из внутриклеточных и внеклеточных источников в сотни-тысячи раз повышает его уровень в цитоплазме. Повышенный уровень Са+ служит активатором ряда процессов, в том числе некоторых протеинкиназ.
На молекулярном уровне передача этого сигнала через мембрану осуществляется цепочкой мембранных белков, последовательно взаимодействующих друг с другом для передачи сигнала малым молекулам, находящимся в цитоплазме. Информация от рецептора на поверхности клетки передается так называемому G-белку, который активирует фермент фосфодиэстеразу, расщепляющую трифосфоинозитид до инозитол – 1,4, 5-трифосфата и 1,2 – диацилглицерина. Инозитолтрифосфат растворим в воде, диффундирует в цитоплазму, где и вызывает описанное выше освобождение кальция. Освободившийся кальций участвует в активации протеинкиназ.
Липофильный диацилглицерин, отличный по своему жирно-кислотному составу от стабильного пула диацилглицеринов, остается в мембране, изменяет ее текучесть и, как уже упоминалось, активирует мембранно-связанную протеинкиназу С,
Эти две различные ветви фосфоинозитидного цикла ведут в конечном счете к фосфорилированию двух различных наборов белков. Оказалось, что с помощью активирующих веществ каждую из ветвей цикла можно привести в действие независимо друг от друга. С другой стороны, применение сочетанного действия фор-боловых эфиров и кальциевых ионофоров помогло установить синергизм двух сигнальных ветвей инозитидного цикла. В таком раздвоенном сигнальном пути совместным действием веществ можно запустить большое число внутриклеточных процессов.
В дальнейшем образовавшиеся 1,2 – диапилглицерин и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается.
7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого позволяет проводить импульс в аксоне с минимальной затратой энергии. Миелиновая оболочка – высокоорганизованная многослойная структура, состоящая из сильно растянутой и модифицированной плазматической мембраны олигодендроглиальной клетки.
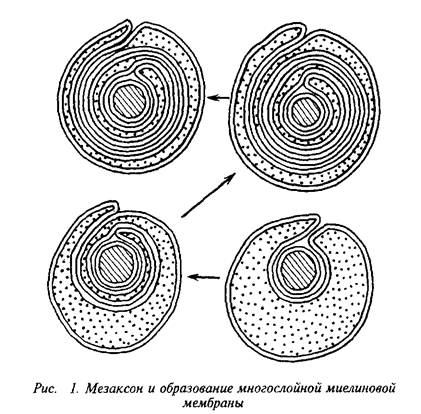
Плазматическая мембрана олигодендроцита образует вокруг аксона сложную мембранную структуру – мезаксон, который является элементарной единицей миелина, имеет пятислойную структуру: белок-липид-белок-липид-белок. Эта пя-тислойная структура, многократно закручиваясь вокруг аксона,