Липиды центральной нервной системы и структура клеточных мембран
Моносиалоганглиозид GM1 хорошо внедряется в мембраны, причем особенно хорошо встраивается молекула GMI, имеющая в своем составе С 2о-эритросфингозин. Возможно, это объясняется его более высокой способностью к мицеллообразованию. Он образует дископодобные мицеллы с М^ЗОО кД, имеющие гидродинамический диаметр около 60 нм.
Интересно, что мицеллы из моносиалоганглиозида GMI по-тенциируют действие ионофора грамицидина D. Ганглиозид-ные мицеллы с заключенными в них молекулами грамицидина включаются в модельную мембрану из фосфатидилсерина и изменяют ее проводимость для ионов калия. После добавления мицелл с ионофором увеличивается время открытия ионных каналов и изменяется их амплитуда.
■ Таким образом, ганглиозидные мицеллы могут участвовать в ионтранспортном процессе в мембране, «маркируя» входы в селективные ионные каналы,
О механизмах и функциональной последовательности действия ганглиозидов известно мало. Встраивание экзогенных ганглиозидов, приводящее к перестройке мембранных ансамблей, изменяет ряд внутриклеточных процессов. Вызванная ганглиозидами дифференциация сопровождается изменением активности Na5K-ATOa3bi, увеличением внутриклеточного уровня цАМФ, уменьшением включения меченого тимидина в ДНК и значительным удлинением фазы G( клеточного цикла. Внедрение ганглиозидов вызывает немедленную перестройку мик-рофиламентной и микротубулиновой системы клеток.
Включение в мембрану экзогенных ганглиозидов усиливает аксональный ток гликозилированных белков и липидов, увеличивает количество гликопротеинов с терминальной манно-зой. Внедрение моносиалоганглиозида GM1 увеличивает в мембране количество эндогенных моносиалоганглиозидов и изменяет активность гликозилтрансфераз: усиливается активность эктофукозилтрансферазы при неизменности активностей сиалил-и галактозилтрансфераз. Внедрение в мембрану трисиалоганг-лиозида GTlb вызывает противоположный эффект.
Недавно выявлено влияние индивидуальных ганглиозидов на фосфорилирование гистона Hj и тубулина, причем в отношении фосфорилирования гистона были особенно эффективны GOJb > GDl* > GTia > GD3> а тубулина – Gxlb > GTla > GQlb > GDla. Показано, что тетрасиалоганглиозид Gglb проявляет зависимое от концентрации влияние на активность Са+ – фос-фолипид-, Са +-кальмодулин-, цАМФ- и цГМФ-активируемых протеиякиназ
8.6 Межклеточное гликозирование ганглиозидов
Своеобразный процесс межклеточного гликозилирования поверхностных гликолипидов и гликопротеинов осуществляется ферментами мембран. Полагают, что гликозилтрансферазы одной клеточной поверхности удлиняют, надстраивают олигосахаридные цепочки гликолипидов и гликопротеинов соседней, противоположной поверхности. Важная регу-ляторная роль в этом процессе принадлежит ионам кальция. Са*+ препятствует образованию субстрат-ферментного комплекса между ганглиозидами и гликозилгрансферазами, а вытеснение его другими ионами способствует межклеточному гликозилированию.
Контактное гликозилирование, как предполагаемый механизм модификации клеточной поверхности в нейрональных мембранах, может быть особенно значимым в образовании синапсов. Вероятно, при этом происходит некая «подгонка» контактирующих мембран.
Роль гликозилирования в синаптической области согласуется с концепцией об участии сиалогликомакромолекул в синаптической передаче и формировании памяти. Полагают, что вхождение сиалогликомакромолекул в контактные зоны является важным звеном молекулярных механизмов в проторении определенных нейрональных путей. Возможно, именно ганглиозиды способствуют образованию ансамблей нейронов, устойчиво связанных друг с другом. Возникновение таких ансамблей исключительно важно для хранения и передачи информации.
8.7 Электрогенность ганглиозидов и ее модификация
Необычайная молекулярная вариабельность ганглиозидов сочетается с лабильной электрогенностью. Для каждой молекулы ганглиозидов характерен свой отрицательный заряд, обусловленный карбоксильной группой сиаловой кислоты. На 1 г ткани мозга приходится не менее 1,3 – Ю анионных групп ганглиозидов. Число анионных групп и, следовательно, уровень отрицательного заряда могут быть объектом регуляции. В этом процессе особая роль принадлежит ферментам – нейрамнни-дазам и сиалилтрансферазам. Они определяют число молекул N-ацетнлнейраминовой кислоты, присутствующих в ганглиозидах, и через цикл сиалирования – десиалирования – отрицательный заряд поверхности.
Сиалилтрансферазы и нейраминидазы находятся на поверхности синаптических мембран там же, где и субстраты, и являются внутренними компонентами синаптической области. В синаптосомалъных мембранах содержится около половиньг ганглиозидов, нейраминидаз и сиалилтрансфераз. Иначе говоря, эти мембраны содержат в 5–6 раз больше ганглиозидов и в 6,5 раз больше нейраминидаз, чем другие плазматические мембраны мозга.
Существенное влияние на поверхностный заряд ганглиозидов в мембране оказывает конформация нейраминовой кислоты и ближайших радикалов. Отщеплению нейраминовой кислоты препятствует соседний N-ацетилгалактозамин. В силу этого гли-козидный кислород нейраминовой кислоты вместе с другими атомами, включающими и карбоксильный кислород N-аиетил-галактозамина, лежит как бы в «кислородной клетке»:
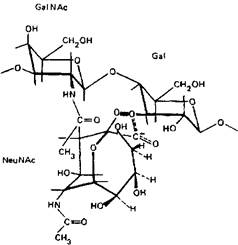
Такая конфигурация атомов вокруг гликозидной связи защищает ее от действия фермента и способствует сохранению отрицательного заряда молекулы. Иная картина наблюдается с ганглиозидами, лишенными N-ацетилгалактозамина: GT3, GD3,
GM3> GM4
Нейраминовая кислота недоступна ферментам, когда карбоксильные группы близлежащих ганглиозидов соединены с Са+:
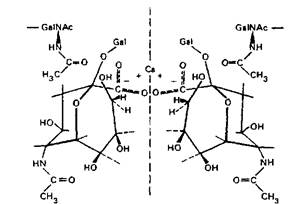
В этом случае исключено не только устранение N-ацетил-нейраминовой кислоты, но и присоединение дополнительного числа ее молекул сиалилтрансферазами.
8.8 Лактонные формы ганглиозидов
Между карбоксильной группой N-апетилнейраминовой кислоты и ее гидроксильными группами могут возникать внутримолекулярные взаимодействия, приводящие к образованию лак-тонов – внутренних сложных эфиров.
В создании лактонов могут участвовать гидроксилы, расположенные'у 4, 7, 8 и 9-го атомов углерода нейраминовой кислоты. Лактоны могут возникать и с участием гидроксильных групп соседней галактозы, приводя к образованию 6-членного кольца:
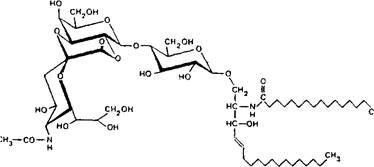
Молекулы нейраминовой кислоты, находящиеся в димерной связи, также образуют лактоны, по структуре аналогичные лактонам коломиновой кислоты, в которой карбоксильная группа одной молекулы связана с гидроксилом 7-го или 9-го атома углерода соседней нейраминовой кислоты.
Лактоны были обнаружены в ганглиозидах мозга. В нейтральной или слабокислой среде терминальная молекула нейраминовой кислоты полисиалоганглиозидов спонтанно образует лак-тон, а в более кислой среде этот процесс затрагивает и другие молекулы нейраминовой кислоты. Установлено, что ионы кальция предотвращают образование лактонов в терминальных молекулах нейраминовой кислоты,