Липиды центральной нервной системы и структура клеточных мембран
Содержание индивидуальных фосфолипидов в коре мозга крысы резко различающихся по своему происхождению и функциям.
В синаптосомах велико содержание жирных кислот – С 22:6, а в миелине высок процент моноеновых кислот – 18:1. Возможно, что высокое содержание докозагексаеновой кислоты в синаптосомах необходимо для активного транспорта ионов, так как активность Na+, К+-АТФазы в них зависит от присутствия полиеновых кислот в фосфолипидах. В мозге имеются регуля-торные механизмы, поддерживающие степень ненасыщенности и специфичность жирнокислотного состава в липидах.
Состав жирных кислот фосфолипидов сннаптосомальных и миеляновых мембран коры мозга обезьяны
|
Шифр жирной кислоты |
Фосфатид ил- | |||||
|
холи н |
этанол амин |
серин+монофосфо-инозитид | ||||
|
синапто-сомы |
миелин |
синапто-сомы |
миелин |
синапто-сомы |
миелин | |
|
16:0 |
50 |
33,1 |
7,4 |
4,9 |
3,5 |
2,6 |
|
18:0 |
12,4 |
17 |
25,5 |
15,9 |
44,3 |
43 |
|
18:1 |
27,2 |
42,3 |
12,1 |
33,2 |
П, 4 |
38,5 |
|
20:1 |
0,7 |
0,9 |
1,6 ■ |
9,3 |
- |
2,5 |
|
20:4 |
3,8 |
3,2 |
10,1 |
11,6 |
8,3 |
6,3 |
|
22:4 |
0,8 |
0,6 |
6,4 |
13,1 |
3,5 |
3,7 |
|
22:6 |
3 |
2,3 |
24,9 |
10,6 |
26,9 |
2,9 |
Изменение жирнокислотного состава приводит к нарушению функциональной деятельности мозга.
1. РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
В мембранах головного мозга имеет место цикл деацилирование – реацилирование, при котором происходит замена жирных кислот в молекуле фосфолипидов, в то время как другие компоненты молекулы остаются неизменными. Этот ацилобменный механизм является особенно важным для включения тех или иных жирных кислот во второе положение остатка глицерина, и его рассматривают как средство локального регулирования физических и функциональных свойств мембран. Существенную роль играет и переход диацильных форм фосфолипидов в моноацильные и обратно. Все это оказывает влияние на такие мембранные процессы, как проницаемость для различных веществ, транспорт ионов и т.д.
Ацилобменные реакции имеют прямое отношение ко многим процессам, влияя на активность ряда ферментов, на синтез простагландинов и чувствительность фоторецепторов. Некоторые исследователи связывают ферментативное деацилирование – реацилирование с эффектом синаптической передачи. Так, под влиянием норадреналина в синаптосомах происходит активирование фосфолипазы А2, отщепляющей жирную кислоту во втором положении глицерофосфолипида. Таким образом, нейромедиатор модифицирует обмен фосфолигавдов в синаптических мембранах путем вовлечения в этот процесс реакций деацилирования. Предложена следующая схема регуляции активности ацилобменного цикла нейромедиаторами.
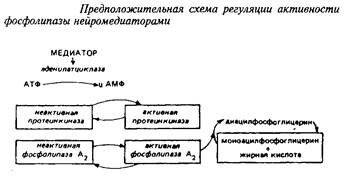
2. ОРГАНИЗАЦИЯ ЛИПИДОВ В МЕМБРАНЕ
Образование липидных молекул в ходе эволюции и выбор именно этих молекул в качестве строительных блоков мембран сыграли решающую роль в возникновении жизни. Липидам принадлежит жизненно важная роль в клетке. Следующие особые физико-химические свойства липидов определяют их роль в построении мембран:
1. Сочетание гидрофильных и липофильных свойств в структуре одной молекулы, их амфифильность.
2. Способность липидов четко ориентироваться на границе раздела фаз, так что полярные группы направлены в водные среды, а неполярные экранированы от них.
3. Способность липидов самопроизвольно упаковываться в прочные, плотные мономолекулярные слои или пленки, устойчивые к сжатию. Плотность такой упаковки зависит от рН, температуры и молекулярной организации липидов. Такие плотные слои создают определенный барьер для диффузии молекул.
4. Способность липидов агрегировать в хорошо упорядоченные сферические, цилиндрические, ламеллярные мицеллы. В мицеллах липиды ориентированы таким образом, чтобы максимальное число полярных групп находилось в контакте с водой, а гидрофобная часть была максимально удалена от контакта с ней.
Способность липидов образовывать прочные мономолекулярные слои лежит в основе молекулярной организации мембран. Более 60 лет назад было высказано предположение, что в основе мембран лежит бимолекулярный слой липидов.
В бимолекулярном липидном слое гидрофобные цепочки молекул липидов направлены друг к другу и внутренность бислоя совершенно гидрофобна, а гидрофильные части образуют поверхности внутреннего и внешнего монослоев, открытые для разнообразного рода взаимодействий.
Липидный состав мембран нервной ткани и распределение липидов по слоям генетически детерминированы. Наружный и внутренний монослои липидов характеризуются планарной и поперечной микрогетерогенностьюу что создает асимметричность мембран. Существует несколько механизмов, поддерживающих асимметричное распределение липидов в мембране. Один из них связан с термодинамической вероятностью размещения липид-ных молекул с учетом их стереоконфигурации, заряда и гидратации полярных групп. Так, основная часть фосфатидилхолина, сфингомиелина, полифосфоинозитидов, холестерина, церебро-зидов и сульфатидов локализована в наружном слое, а амино-фосфолипиды находятся во внутреннем, цитоплазматическом слое. Неодинакова степень ненасыщенности монослоев: во внутреннем обнаруживается 2/3 двойных связей в жирных кислотах липидов, а в наружном – только 1/3.