Страница
7
Второй этап связан с регенерацией исходного метаболита — глюкозо-6-фосфата. Из рибулозо-5-фосфата под действием эпимеразы (4) образуется ксилулозо-5-фосфат, а под действием изомеразы (5) — рибозо-5-фосфат. Рекомбинации сахаров с участием транскетолазы (6,8) и трансальдолазы (7) приводят к появлению 3-ФГА и седогептулозо-7-фосфата, затем эритрозо-4-фосфата (7) и фруктозо-6-фосфата(8); в результате образуются фруктозо-6- фосфаты, которые изомеризуются в глюкозо-6-фосфат (12).
Как видно из рис.5, 6 молекул глюкозо-6-фосфата, участвуя в ПФП дыхания, дают 6 молекул рибулозо-5-фосфата и 6С02, после чего из 6 молекул рибулозо-5-фосфата регенерируют 5 молекул глюкозо-6-фосфата. Для каждого оборота цикла суммарное уравнение ПФП имеет следующий вид:
6 Глюкозо-6-фосфат + 12НАДФ + + 7Н20 -->5Глюкозо-6-фосфат + 6С02 + 12НАДФH + 12Н+ + Н3Р04
Энергетический выход ПФП и его роль в обмене веществ.
Универсальным донором водорода для электронтранспортной цепи дыхания служит НАДH, содержание которого в растительных тканях всегда значительно выше, чем НАДФH. В нормальных условиях НАДФ+ находится в клетках в восстановленной форме НАДФH, тогда как НАД+ — в окисленной. Доказано, что НАДФH окисляется медленнее, чем НАДH. Если при окислении субстрата образуется НАДФH, как, например, при апотомическом окислении глюкозо-6-фосфата, то атомы водорода перед поступлением в электронтранспортную цепь должны быть переданы на. НАД+ (трансгидрогеназцая реакция). Если бы все 12 пар протонов от НАДФH, которые образуются при полном окислении молекулы глюкозо-6-фосфата по ПФП, были бы переданы через ЭТЦ на 02, то получилось бы 3 АТФ х 12 = 36 АТФ, что составляет 41,868 кДж х х 36 = 1507 кДж/моль. Практически это не уступает энергетическому выходу дихотомического пути дыхания (гликолиз и цикл Кребса), в котором образуется 1591 кДж/моль (38 АТФ,)1
Однако основное назначение ПФП состоит в участии не столько в энергетическом, сколько в пластическом обмене клеток. Это участие в пластическом обмене включает несколько аспектов:
1. НАДФH используется главным образом в различных синтетических реакциях.
2.В ходе пентозофосфатного цикла синтезируются пентозы, входящие в состав нуклеиновых кислот и различных нуклеотидов (пиридиновых, флавиновых, адениловых и др.). Для животных и других гетеротрофных организмов ПФП — единственный способ образования пентоз (рибоз и дезоксирибоз) в клетке. Рибозы необходимы для синтеза АТФ, GTP, UTP и других нуклеотидов. Коферменты НАД+, НАДФ+, ФАД, коэнзим А — тоже нуклеотиды и в их состав входит рибоза.
3.ПФП имеет большое значение как источник образования углеводов с различным числом углеродных атомов в цепи (от С3 до Су). Эритрозо-4-фосфат, возникающий в ПФП, необходим для синтеза шикимовой кислоты — предшественника многих ароматических соединений, таких, как ароматические аминокислоты, витамины, дубильные и ростовые вещества, лигнин клеточных стенок и др.
4.Компоненты ПФП (рибулозо-1,5-дифосфат, НАДФH) принимают участие в темновой фиксации С02. По существу, ПФП представляет собой обращенный фотосинтетический (восстановительный) цикл Кальвина. Только две из 15 реакций цикла Кальвина специфичны для фотосинтеза, остальные участвуют в окислительном ПФП дыхания и гликолизе.
В хлоропластах окислительный ПФП функционирует в темноте, предотвращая резкое изменение концентрации НАДФH в отсутствие света. Кроме того, триозофосфаты этого цикла в хлоропластах превращаются в 3-ФГК, что важно для поддержания в них уровня АТФ в темноте.
Окисление глюкозы по ПФП осуществляется в результате 12 реакций, тогда как в дихотомический (гликолитический) путь через ПВК и далее через цикл ди- и трикарбоновых кислот включаются более 30 различных реакций.
Не следует, однако, думать, что окисление глюкозо-6-фосфата по схеме, представленной на рис. 5, идет во всех клетках до конца. Очень часто на одном из этапов ПФП переходит в гликолитический. Таким этапом может быть, в частности, транскетолазная реакция (рис. 5, реакция 8), в результате которой ксилулозо-5-фосфат и эритрозо-4-фосфат превращаются во фруктозо-6-фосфат и 3-ФГА — субстраты, общие для гликолиза и ПФП.
4.3 Глиоксилатный цикл. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Этот цикл в 1957 г. был впервые описан у бактерий и плесневых грибов Г. Л. Корнбергом и Г. А. Кребсом. Затем оказалось; что он активно функционирует в про- растающих семенах масличных растений и в других растительных объектах, где запасные жиры превращаются в сахара (глюконеогенез). Глиоксилатный цикл локализован не в митохондриях, как цикл Кребса, а в специализированных микротелах — глиоксисомах. В клетках животных этот цикл отсутствует.
В глиоксилатном цикле из ЩУК и ацетил-СоА синтезируется лимонная кислота, образуются цис-аконитовая и изолимонная (изоцитрат), как и в цикле Кребса . Затем изолимонная кислота под действием изоцитрат-лиазы распадается на глиоксиловую и янтарную кислоты. Глиоксилат с участием малатсинтазы взаимодействует со второй молекулой ацетил-СоА, в результате чего синтезируется яблочная кислота, которая окисляется до ЩУК.
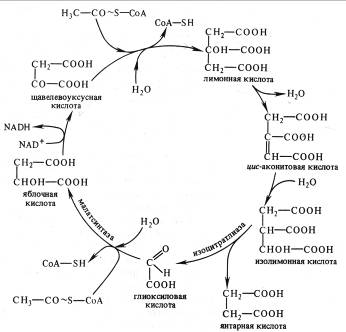
Рис. 6. Глиоксилатный цикл
Таким образом, в отличие от цикла Кребса в глиоксилатном цикле в каждом обороте участвует не одна, а две молекулы ацетил-СоА и этот активированный ацетил используется не для окисления, а для синтеза янтарной кислоты. Янтарная кислота выходит из глиоксисом, превращается в ЩУК и участвует, в глюконеогенезе (обращенном гликолизе) и других процессах биосинтеза. Глиоксилатный цикл позволяет утилизировать запасные жиры, при распаде которых образуются молекулы ацетил-СоА.
Регуляция ПФП и глиоксилатного цикла. Пентозофосфатный путь окисления регулируется концентрацией НАДФ + , так как содержит две НАДФ-зависимые дегидрогеназы (глюкозо-6- фосфатдегидрогеназу и 6-фосфоглюконатдегидрогеназу). Он регулируется также уровнем синтезов в клетке, потребляющих НАДФH (например, синтезом аминокислот и белков). Их высокий уровень приводит к увеличению содержания окисленного НАДФ + , что стимулирует ПФП.
В регуляции соотношения между ПФП и гликолизом принимает участие ряд интермедиатов: неорганический фосфат, 6-фосфоглюконовая кислота, эритрозо-4-фосфат. Недостаток неорганического фосфата подавляет гликолиз и активирует ПФП. 6-Фосфоглюконовая кислота служит ингибитором гликолитического фермента фосфофруктокиназы (глюкозофосфатизомеразы), что способствует функционированию ПФП. Эритрозо-4-фосфат, являясь субстратом транскетолазной и трансальдолазной реакций, может тормозить активность ферментов гликолиза и тем самым переключать превращения углеводов с гликолитического на пентозофосфатный путь.
Активность глиоксилатного цикла снижается при повышении концентрации оксалоацетата, который ингибирует сукцинатдегидрогеназу ЦТК. Другой ингибитор цикла — фосфоенолпируват подавляет активность изоцитратлиазы.
5. Цепь переноса водорода и электрона (дыхательная цепь). Комплексы переноса электронов. Окислительное фосфорилирование. Хемиосмотическая теория окисления и фосфорилирования. Механизмы сопряжения процесса транспорта электронов с образованием АТФ