Страница
6
Следует отметить, что большинство реакций цикла обратимы, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзергонические реакции — цитратсинтазная и сукцинил-СоА-синтетазная.
На протяжении одного оборота цикла при окислении пирувата происходит выделение трех молекул С02, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль Н2О в цикле Кребса подтверждает правильность уравнения Палладина, который постулировал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям — коферментов дегидрогеназ) переносится на кислород .
Выше отмечалось, что цикл Кребса был открыт на животных объектах. Существование его у растений впервые доказал английский исследователь А. Чибнелл (1939). В растительных тканях содержатся все кислоты, участвующие в цикле; обнаружены все ферменты, катализирующие превращение этих кислот; показано, что малонат — ингибитор сункцинатдегидрогеназы — тормозит окисление пирувата и резко снижает поглощение 02 в процессах дыхания у растений. Большинство ферментов цикла Кребса локализовано в мАТФиксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрии.
Энергетический выход цикла Кребса, его связь с азотным обменом. Цикл Кребса. играет чрезвычайно важную роль в обмене веществ растительного организма. Он служит конечным этапом окисления не только углеводов, но также белков, жиров и других соединений. В ходе реакций цикла освобождается основное количество энергии, содержащейся в окисляемом субстрате, причем большая часть этой энергии не теряется для организма, а утилизируется при образовании высокоэнергетических конечных фосфатных связей АТФ.
Каков же энергетический выход цикла Кребса? В ходе окисления пирувата имеют место 5 дегидрирований, при этом получаются 3НАДH, НАДФH (в случае изоцитратдегидрогеназы) и ФАДH2. Окисление каждой молекулы НАДH (НАДФH) при участии компонентов электронтранспортной цепи митохондрий дает по 3 молекулы АТФ, а окисление ФАДH2 — 2АТФ. Таким образом при полном окислении пирувата образуются 14 молекул АТФ. Кроме того, 1 молекула АТФ синтезируется ; в цикле Кребса в ходе субстратного фосфорилирования. Следовательно, при окислении одной молекулы пирувата может образоваться 15 молекул АТФ. А поскольку в процессе гликолиза из молекулы глюкозы возникают две молекулы пирувата, их окисление даст 30 молекул АТФ.
Итак, при окислении глюкозы в процессе дыхания при функционировании гликолиза и цикла Кребса в общей сложности образуются 38 молекул АТФ (8 АТФ связаны с глико- лизом). Если принять, что энергия третьей сложноэфирнои фосфатной связи АТФ равняется 41,87 кДж/моль (10 ккал/моль), то энергетический выход гликолитического пути аэробного дыхания составляет 1591 кДж/моль (380 ккал/моль).
Значение цикла Кребса не ограничивается его вкладом в энергетический обмен клетки. Не менее важную роль играет то обстоятельство, что многие промежуточные продукты цикла используются при синтезе различных соединений. Из кетокислот в ходе реакций переаминирования образуются аминокислоты. Для синтеза липидов, полиизопренов, углеводов и ряда других соединений используется ацетил-СоА.
Регуляция цикла Кребса. Дальнейшее использование образующегося из пирувата ацетил-СоА зависит от энергетического состояния клетки. При малой энергетической потребности клетки дыхательным контролем тормозится работа дыхательной цепи, а следовательно, реакций ЦТК и образования интермедиатов цикла, в том числе оксалоацетата, вовлекающего ацетил-СоА в цикл Кребса. Это приводит к большему использованию ацетил-СоА в синтетических процессах, которые также потребляют энергию.
Особенностью регуляции ЦТК является зависимость всех четырех дегидрогеназ цикла (изоцитратдегидрогеназы, α-кетоглутаратдегидрогеназы, сукцинатдегидрогеназы, малатдегидрогеназы) от отношения [НАДH]/[НАД+]. Активность цитратсинтазы тормозится высокой концентрацией АТФ и собственным продуктом — цитратом. Изоцитратдегидрогеназа ингибируется НАДH и активируется цитратом. α-Кето- глутаратдегидрогеназа подавляется продуктом реакции — сукцинил-СоА и активируется аденилатами. Окисление сукцината сукцинатдегидрогеназой тормозится оксалоацетатом и ускоряется АТФ, АДФ и восстановленным убихиноном (QH2). Наконец, малатдегидрогеназа ингибируется оксалоацетатом и у ряда объектов — высоким уровнем АТФ. Однако степень участия величины энергетического заряда, или уровня адениновых нуклеотидов, в регуляции активности цикла Кребса у растений до конца не выяснена.
Регулирующую роль может играть также альтернативный путь транспорта электронов в растительных митохондриях. В условиях высокого содержания АТФ, когда активность основной дыхательной цепи снижена, окисление субстратов через альтернативную оксидазу (без образования АТФ) продолжается, что поддерживает на низком уровне отношение НАДH/НАД+ и снижает уровень АТФ. Все это позволяет циклу Кребса функционировать.
4.2 Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
В клетках растений наряду с гликолизом и циклом Кребса, являющимся главным поставщиком свободной энергии в процессах дыхания, существует и другой важнейший способ катаболизма гексоз — пентозофосфатный путь (ПФП), в котором участвуют пятиуглеродные сахара (пентозы). Этот путь дыхания известен также как гексозомонофосфатный цикл, пентозный шунт или апотомическое окисление. Окисление глюкозы (глюкозо-6-фосфата) по этому пути связано с отщеплением первого (альдегидного) атома углерода в виде С02 (отсюда и название — апотомический путь).
Пентозофосфатный путь дыхания открыт в 1935—1938 гг. в результате исследований О. Варбурга, Ф. Диккенса, В. А. Энгельгардта и позднее Ф. Липмана. Установлено, что все реакции ПФП протекают в растворимой части цитоплазмы клетки, а также в пропластидах и хлоропластах. ПФП дыхания особенно активен в тех клетках и тканях растений, в которых интенсивно идут синтетические процессы, такие, как синтез липидных компонентов мембран, нуклеиновых кислот, клеточных стенок, фенольных соединений.
В ПФП АТФ используется только для образования исходного продукта. ПФП, как и цикл Кребса, — циклический процесс, поскольку окисление глюкозы сопровождается регенерацией исходного субстрата ПФП — глюкозо-6-фосфата.
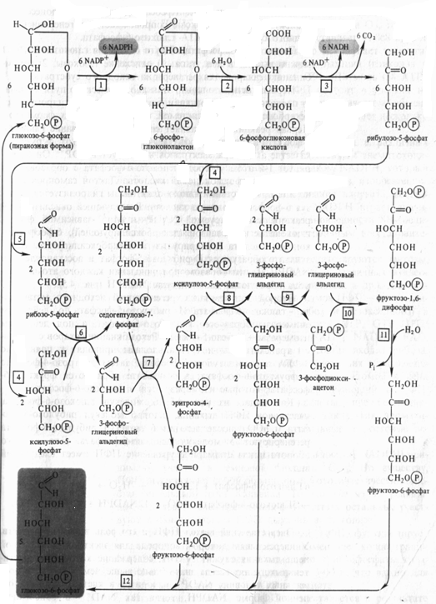
Рис. 5. Пентозофосфатный цикл
Этапы пентозофосфатного пути окисления глюкозы. В ПФП можно выделить два этапа: 1) окисление глюкозы, 2) рекомбинацию Сахаров для регенерации исходного субстрата.
Первый, окислительный, этап апотомического пути включает последовательные реакции, катализируемые дегидрогеназнодекарбоксилрующей системой, состоящей из трех ферментов. Первая реакция представляет собой Дегидрирование глюкозо- 6-фосфата глюкозо-6-фосфатдегидрогеназой (1). Этот фермент в качестве акцептора электронов использует НАДФ+. Он дегидрирует 1-й атом углерода глюкозо-6-фосфата с образованием лактона 6-фосфоглюконовой кислоты. Лактон самопроизвольно или под действием глюконолактоназы (2) гидролизуется, образуя 6-фосфоглюконовую кислоту. В следующей окислительной реакции, катализируемой НАДФ- и Мп2 +-зависимой фосфо- глюконатдегидрогеназой (3) (декарбоксилирующей), 6-фосфоглюконовая кислота дегидрируется и декарбоксилируется. В результате образуются рибулозо-5-фосфат и восстановленный НАДФH. Таким образом, при окислении каждого атома углерода образуются две молекулы НАДФH (рис. 5).