Дыхание растений
В растущих клетках митохондриальный матрикс становится менее плотным, количество крист растет — это коррелирует с увеличением интенсивности дыхания. В процессе дыхания ультраструктура митохондрий меняется. В том случае, если в митохондриях протекает активный процесс преобразования энергии окисления в энергию АТФ, внутренняя часть митохондрий становится более компактной.
Митохондрии имеют свой онтогенез. В меристематических клетках можно наблюдать инициальные частицы, которые представляют собой округлые образования, окруженные двойной мембраной. Диаметр таких инициальных частиц составляет 50 нм. По мере роста клетки инициальные частицы увеличиваются в размере, удлиняются и их внутренняя мембрана образует выросты, перпендикулярные оси митохондрий. Вначале образуются промитохондрии. Они еще не достигают окончательного размера и имеют мало крист. Из промитохондрий образуются митохондрии. Сформировавшиеся митохондрии делятся путем перетяжки или почкованием. Свойства митохондрий (белки, структура) закодированы частично в ДНК митохондрий, а частично в ядре. Сопоставление размеров митохондриальной ДНК с числом и размером митохондриальных белков показывает, что в ней заложено информации почти для половины белков. Это и позволяет считать митохондрии полуавтономными, т. е. не полностью зависящими от ядра. Они имеют собственную ДНК и собственную белоксинтезирующую систему, и именно с ними и с пластидами связана так называемая цитоплазматическая наследственность. В большинстве случаев это наследование по материнской линии, так как инициальные частицы митохондрий локализованы в яйцеклетке. Таким образом, митохондрии всегда от митохондрий.
Широко обсуждается вопрос, как рассматривать митохондрии и хлоропласты с эволюционной точки зрения. Еще в 1921 г. русский ботаник Б. М. Козо-Полянский высказал мнение, что клетка — это симбиотрофная система, в которой сожительствует несколько организмов. В настоящее время эта гипотеза имеет много сторонников. Согласно гипотезе симбиогенеза, митохондрии — это в прошлом самостоятельные организмы. По мнению Марголис, это могли быть эубактерии, содержащие ряд дыхательных ферментов. На определенном этапе эволюции они внедрились в примитивную содержащую ядро клетку. Оказалось, что ДНК митохондрий и хлоропластов по своей структуре резко отличается от ядерной ДНК высших растений и сходна с бактериальной ДНК (кольцевое строение). Сходство обнаруживается и в величине рибосом. Однако доказательств еще недостаточно и окончательного вывода по этому вопросу пока сделать невозможно.
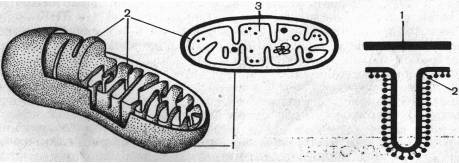
1- наружная мембрана, 2- внутренняя мембрана, 3- матрикс.
Рис. 10. Схема строения митохондрий
8. Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Соссюр, работая с зелеными растениями в темноте, обнаружил, что они выделяют С02 даже в бескислородной среде. Л. Пастер нашел, что в темноте в отсутствие кислорода в растительных тканях наряду с выделением С02 образуется спирт, т. е. идет спиртовое брожение. Он пришел к выводу, что в растительных тканях, так же как и у бактерий, возможно спиртовое брожение.
Немецкий физиолог Э. Ф. Пфлюгер (1875), изучая дыхание животных объектов, показал, что лягушки, помещенные в среду без кислорода, некоторое время остаются живыми и при этом выделяют С02. Пфлюгер назвал это дыхание интрамолекулярным, т. е. дыханием за счет внутримолекулярного окисления субстрата. Предполагалось, что интрамолекулярное дыхание — начальный этап нормального аэробного дыхания. Эту точку зрения поддержал Б. Пфеффер — немецкий физиолог растений, который распространил ее на растительные организмы. На основе этих работ Пфеффером и Пфлюгером были предложены следующие два уравнения, описывающие механизм дыхания:
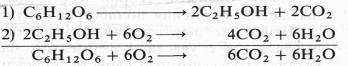
На первом, анаэробном, этапе происходит спиртовое брожение, образуются две молекулы этанола и две молекулы С02. Затем в присутствии кислорода спирт, взаимодействуя с ним, окисляется до С02 и Н20.
С. П. Костычев (1910) пришел к выводу, что это уравнение не соответствует действительности. Он экспериментально доказал, что этанол не может быть промежуточным продуктом нормального аэробного дыхания у растений по двум причинам: во-первых, он ядовит для растений и не может накапливаться, во-вторых, этанол окисляется растительными тканями значительно хуже, чем глюкоза. Костычев предложил свою формулу связи анаэробной и аэробной частей дыхания и различных видов брожения.
В опытах Костычева и его сотрудников (1912—1928) было показано, что если растительные ткани кратковременно выдержать в бескислородной среде, а затем дать кислород, то наблюдается резкое усиление дыхания, т.е. в ходе анаэробной фазы накапливаются промежуточные продукты, которые в присутствии кислорода быстро используются. Ингибиторы, блокирующие брожение, например NaF, блокируют и аэробное дыхание. Ингибиторный анализ (применение ингибиторов специфического действия), выделение и идентификация продуктов окислительного распада глюкозы привели Костычева к выводу о том, что промежуточным продуктом может быть уксусный альдегид. Благодаря работам немецкого биохимика К. Нейберга, Костычева и других стало очевидным, что дыхание и все виды брожения связаны между собой через пировиноградную кислоту (ПВК):
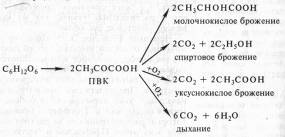
Таким образом, теория Костычева о генетической связи дыхания и брожения полностью подтвердилась.
Взаимосвязь дыхания с другими процессами обмена.
Дыхание тесно связано с другими процессами метаболизма.Необходимо подчеркнуть, что, несмотря на противоположную направленность двух центральных процессов растительного организма —.фотосинтеза и дыхания и на их рассредоточенность в разных органеллах клетки, между ними существует тесная взаимосвязь. Прежде всего для протекания процесса дыхания органические вещества (субстраты). Таким субстратами в первую очередь являются углеводы, которые у зеленых растений образуются в процессе фотосинтеза. Вместе с тем превращение веществ в процессе фотосинтеза и дыхания идет через ряд сходных промежуточных продуктов. Особенно много сходного в превращениях между фотосинтетическим циклом Кальвина и реакциями пентозофосфатного пути дыхательного обмена. Как в том, так и в другом случае происходят взаимные превращения Сахаров с разной длиной углеродной цепочки (3, 4, 5, 6 и 7 углеродных атомов). По-видимому, несмотря на различное распределение (компартментацию) этих метаболитов в клетке, между ними существует обмен. Иначе го вор я, промежуточные. продукты дыхания могут быть использованы в процессе фотосинтеза. Одновременно возможен и обратный процесс. Много общего в энергетике фотосинтеза и дыхания в процессах фотосинтетического и окислительного фосфорилирования. Между этими двумя процессами возможен обмен энергетическими эквивалентами. АТФ, образовавшаяся на свету при фотосинтетическом фосфорилировании, может служить основным источником энергии для различных биосинтетических процессов, заменяя АТФ, образовавшуюся в процессе дыхания. С другой стороны, АТФ и НАДФ-Н, образовавшиеся в процессе дыхания, могут быть использованы для реакций цикла Кальвина. Имеются наблюдения, что на свету основными органеллами, поставляющими АТФ, являются хлоропласты.