Страница
4
Английский биохимик А. Гарден и ученик К. А. Тимирязева Л. А. Иванов в 1905 г. независимо показали, что в процессе спиртового брожения наблюдается связывание неорганического фосфата и превращение его в органическую форму. Гарден установил, что глюкоза подвергается анаэробному распаду только после ее фосфорилирования. Полностью весь процесс гликолиза расшифровали немецкие биохимики Г. Эмбден, О. Ф. Мейергоф и советский биохимик Я. О. Парнас, с именами которых связывают название этого катаболического
Цепь реакций, составляющих суть гликолиза, можно разбить на три этапа:
I. Подготовительный этап — фосфорилирование гексозы и ее расщепление на две фосфотриозы.
II. Первое субстратное фосфорилирование, которое начинается с 3-фосфоглицеринового альдегида и кончается 3-фосфоглицериновой кислотой. Окисление альдегида до кислоты связано с освобождением энергии. В этом процессе на каждую фосфотриозу синтезируется одна молекула АТФ.
III. Второе субстратное фосфорилирование, при котором 3-фосфоглицериновая кислота за счет внутримолекулярного окисления отдает фосфат с образованием АТФ.
Поскольку глюкоза стабильное соединение, на ее активацию необходима затрата энергии, что осуществляется в процессе образования фосфорных эфиров глюкозы в ряде подготовительных реакций. Глюкоза (в пиранозной форме) фосфорилируется АТФ с участием гексокиназы (1), превращаясь в глюкозо-6-фосфат, который изомеризуется в фруктозо-6-фосфат с помощью глюкозофосфатизомеразы (2).
Этот - переход необходим для образования более лабильной фуранозной формы молекулы гексозы. Фруктозо-6-фосфат фосфорилируется вторично фосфофрукгокиназой с использованием еще одной молекулы АТФ (3).
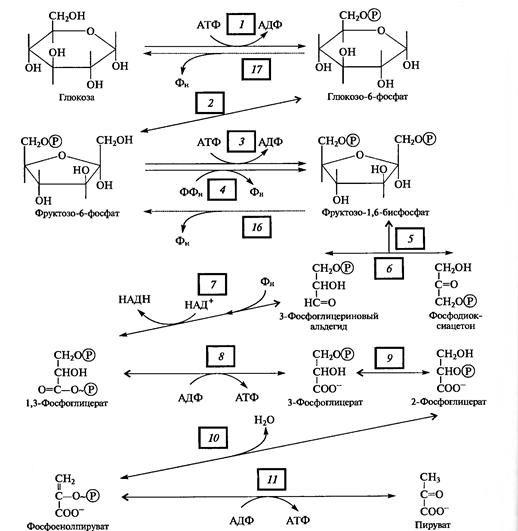
Рис. 2. Реакции гликолиза
Фруктозо-1,6-дифосфат — лабильная фуранозная форма с симметрично расположенными фосфатными группами. Обе эти группы несут отрицательный заряд, отталкиваясь друг от друга электростатически. Такая структура легко расщепляется альдолазой на две фосфотриозы. Следовательно, смысл подготовительного этапа состоит в активации молекулы гексозы за счет двойного фосфорилирования и перевода в фуранозную форму с последующим распадом на 3-фосфоглицериновый альдегид (3-ФГА) и фосфодиоксиацетон (ФДА) (5), причем бывший 6-й атом углерода в молекуле глюкозы и фруктозы (фосфорилированный) становится 3-м в 3-ФГК, а 1-й атом углерода фруктозо-1,6-дифосфата остается 1-м углеродом (фосфорилированным) в ФДА. 3-ФГА и ФДА легко превращаются друг в друга с участием триозофосфатизомеразы (6). Из-за расщепления молекулы гексозы на две триозы гликолиз иногда называют дихотомическим путем окисления глюкозы.
С 3-ФГА начинается II этап гликолиза — первое субстратное фосфорилирование. Фермент дегидрогеназа фосфоглицеринового альдегида (НАД-зависимый SH-фермент) (7) образует с 3-ФГА фермент-субстратный комплекс, в котором происходит окисление субстрата и передача электронов и протонов на НАД+. В ходе окисления фосфоглицеринового альдегида до фосфоглицериновой кислоты в фермент-субстратном комплексе возникает меркаптанная высокоэнергетическая связь (т. е. связь с очень высокой свободной энергией гидролиза). Далее осуществляется фосфоролиз этой связи, в результате чего SH-фермент отщепляется от субстрата, а к остатку карбоксильной группы субстрата присоединяется неорганический фосфат, причем ацилфосфатная связь сохраняет значительный запас энергии, освободившейся в результате окисления 3-ФГА. Высокоэнергетическая фосфатная группа с помощью фосфоглицераткиназы передается на АДФ и образуется АТФ (8). Так как в данном случае высокоэнергетическая ковалентная связь фосфата формируется прямо на окисляемом субстрате, такой процесс получил название субстратного фосфорилирования. Таким образом, в результате II этапа гликолиза образуются АТФ и восстановленный НАДH.
Последний этап гликолиза — второе субстратное фосфорилирование. З-Фосфоглицериновая кислота с помощью фосфоглицератмутазы превращается в 2-фосфоглицериновую кислоту (9). Далее фермент енолаза катализирует отщепление молекулы воды от 2-фосфоглицериновой кислоты (10). Эта реакция сопровождается перераспределением энергии в молекуле, в результате чего образуется фосфоенолпируват — соединение, содержащее высокоэнергетическую фосфатную связь. Таким образом, в этом случае высокоэнергетическая фосфатная связь формируется на основе того фосфата, который имелся в самом субстрате. Этот фосфат при участии пируваткиназы (11) передается на АДФ и образуется АТФ, а енолпируват самопроизвольно переходит в более стабильную форму — пируват — конечный продукт гликолиза.
Энергетический выход гликолиза. При окислении одной молекулы глюкозы образуются две молекулы пировиноградной кислоты. При этом за счет первого и второго субстратного фосфорилирования образуются четыре молекулы АТФ. Однако две молекулы АТФ тратятся на фосфорилирование гексозы на I этапе гликолиза. Таким образом, чистый выход гликолитического субстратного фосфорилирования составляет две молекулы АТФ.
Кроме того, на II этапе гликолиза на каждую из двух молекул фосфотриоз восстанавливается по одной молекуле НАДH. Окисление одной молекулы НАДH в электронтранспортной цепи митохондрий в присутствии 02 сопряжено с синтезом трех молекул АТФ, а в расчете на две триозы (т. е. на одну молекулу глюкозы) - шесть молекул АТФ. Таким образом, всего в процессе гликолиза (при условии последующего окисления НАДН) образуются восемь молекул АТФ. Поскольку свободная энергия гидролиза одной молекулы АТФ во внутриклеточных условиях составляет около 41,868 кДж/моль (10 ккал), восемь молекул АТФ дают 335 кДж/моль, или 80 ккал. Таков полный энергетический выход гликолиза в аэробных условиях.
Обращение гликолиза. Возможность обращения гликолиза определяется обратимостью действия большинства ферментов, катализирующих его реакции. Однако реакции фосфорилирования глюкозы и фруктозы, а также реакция образования пировиноградной кислоты из фосфоенолпирувата, осуществляемые с помощью киназ, необратимы. На этих участках процесс обращения идет благодаря использованию обходных путей. Там, где функционируют гексокиназа и фруктокиназа, происходит дефосфорилирование — отщепление фосфатных групп фосфатазами.
Превращение пирувата в фосфоенолпируват также не может осуществиться путем прямого обращения пируваткиназной реакции вследствие большого перепада энергии. Первая реакция обращения гликолиза на этом участке катализируется митохондриальной пируваткарбоксилазой в присутствии АТФ и ацетил-СоА (последний выполняет функции активатора). Образующаяся щавелевоуксусная кислота (ЩУК), или оксалоацетат, восстанавливается затем в митохондриях до малата при участии НАД-зависимой малатдегидрогеназы (МДГ). Затем малат транспортируется из митохондрий в цитоплазму, где окисляется НАД-зависимой цитоплазматической малатдегидрогеназой снова до ЩУК. Далее под действием ФЕП-карбоксикиназы из оксалоацетата образуется фосфоенолпируват. Фосфорилирование в этой реакции осуществляется за счет АТФ.