Фотосинтез как основа энергетики биосферы
Передача (миграция) энергии по пигментам антенных комплексов происходит по принципу индуктивного резонанса (без флуоресценции и переноса заряда). Природа индуктивного резонанса сострил в следующем. Каждая молекула хлорофилла, поглотившая квант света и перешедшая в синглетное возбужденное состояние, является молекулярным осциллятором. Возникающее вокруг возбужденной молекулы переменное электрическое поле с определенной частотой колебаний индуцирует осцилляцию диполя (электрон — ядро) соседней молекулы. При этом молекула-донор переходит в основное состояние, а молекула-акцептор — в возбужденное. Условиями для резонансного переноса энергии электронного возбуждения служат малые расстояния между молекулами, не превышающие 10 нм, и перекрытие частот колебаний у двух взаимодействующих молекул.
Флуоресценция каждой молекулы-донора имеет более длинноволновый максимум по сравнению с максимумом ее поглощения (в соответствии с правилом Стокса) и в большей или меньшей степени перекрывает зону поглощения акцепторной молекулы. Миграция энергии осуществляется от коротковолновых пигментов в сторону все более длинноволновых, т. е. пигментов с более низким уровнем синглетного возбужденного состояния.
В антенных комплексах перенос энергии осуществляется в ряду: каротин (400-550 нм)--► хлорофилл b (650 нм)--►хлорофиллы a (660-675 нм)---► П680 (ФС II). Скорость резонансного переноса энергии от молекулы к молекуле 10-10 — 10-9 с, причем эффективность переноса между молекулами хлорофилла достигает 100, а между молекулами каротина и хлорофилла — лишь 40%.
Реакционные центры. Белки, содержащие длинноволновые формы хлорофиллов (П680 в ФС II и П700 в ФС I) в количестве одна молекула на 200 — 400 молекул других хлорофиллов и способные к первичному фотохимическому разделению зарядов, называют реакционными центрами.
Первичное разделение зарядов в реакционных центрах происходит между молекулами хлорофилловой природы и связано с транспортом электронов. Функцию первичного донора электронов выполняет молекула хлорофилла, находящаяся в синглетном возбужденном состоянии. В реакционном центре ФС II, перешнуровывающем мембрану, первичным донором электронов служит П680, а первичным акцептором — феофетин. В реакционном центре ФС I первичный донор электронов— П700, а акцептор — мономерная форма хлорофилла а695 (А1).
Порядок реакций в реакционном центре ФС II определяется продолжительностью того или иного состояния молекул. Молекулы находятся в синглетном возбужденном состоянии 10-10 — 10-9 с. Первичные акцепторы (Фф или А1) получают электроны от своих доноров (П680 или П700 соответственно) за время около 10-12 с. Время обратных рекомбинаций от Фф- на П680+ и от A1- на П700+ во много раз больше (10-6 с). То же самое относится к последующим парам взаимодействующих молекул. Поэтому весь ход реакций резко сдвинут вправо. В реакционных центрах энергия возбужденного состояния хлорофилла преобразуется в энергию разделенных зарядов, т. е. превращается в химическую энергию.
5.2 Фотохимический этап. Электронно-транспортная цепь фотосинтеза. Представления о функционировании двух фотосистем. Фотофосфорилирование. Системы фотоокисления воды и выделения кислорода при фотосинтезе. Связь фотосинтетической ассимиляции С02 с фотохимическими реакциями
Нециклический и циклический транспорт электронов.
Для восстановления одной молекулы NADP+ в процессе фотосинтеза необходимы два электрона и два протона, причем донором электронов является вода. Фотоиндуцированное окисление воды осуществляет ФС II, восстановление NADP+ — фотосистема I. Следовательно, эти две фотосистемы должны функционировать во взаимодействии.
Исходя из данных о квантовом выходе фотосинтеза (необходимы 8 квантов света для выделения одной молекулы 02) из состава компонентов, входящих в ФС I и ФС II, и величин их окислительно-восстановительного потенциала была разработана схема последовательности реакций в световой фазе фотосинтеза.
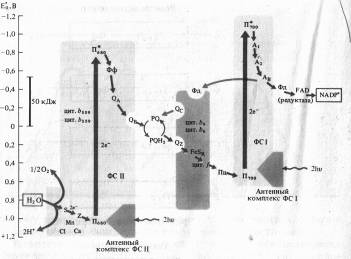
Из-за сходства с буквой Z эта схема, получила название Z-схемы. Впервые принцип Z-схемы был предложен Р. Хиллом и Ф. Бендаллом (1960) и экспериментально подтвержден работами Л. Дюйзенса (1961). В настоящее время Z-схема, или схема нециклического транспорта электронов при фотосинтезе является общепризнанной и непрерывно пополняется новыми деталями.
В ФС II димер П680, поглотив энергию, эквивалентную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, отдает 2 электрона феофетину (Фф). От Фф электроны, теряя энергию, последовательно передаются на пластохиноны ФС 11 - QA и QB, на пул липидорастворимых молекул пластохинона (PQ), переносящих через липидную фазу мембраны электроны и протоны, на железосерный белок FeSK и цитохром цитохромного комплекса b6 — f, восстанавливая Cu-содержащий белок пластоцианин (Пц).
Вакантные места («дырки») в Пб80 заполняются двумя электронами из содержащего Мп переносчика электронов Z, который в свою очередь восстанавливается с участием системы S. Окисленный белковый комплекс S связывает воду и восстанавливается за счет электронов воды. Для осуществления этой реакции в белковом комплексе S необходимы Мп и С1-, а также Са2 + .
Однако описанная цепь реакций резко замедляется, если не возбуждена ФС I, так как в этом случае весь Пц переходит в восстановленное состояние. При возбуждении П700 в реакционном центре ФС I энергией, эквивалентной 2 квантам длинноволнового красного света, 2 электрона захватываются мономерной формой хлорофилла a (A1) и затем последовательно передаются переносчикам электронов А2 и Ав (железосерные белки FeS), ферредоксину (водорастворимый FeS-белок) и ферредоксин: NADP-оксидоредуктазе с FAD в качестве кофактора. Наконец, редуктаза восстанавливает NADP + .
На вакантные места в П700+ переходят электроны с Пц, и нециклическая цепь переноса электронов таким образом замыкается. Z-схема убедительно объясняет «эффект усиления Эмерсона», так как только при совместном использовании коротковолновых (ФС II) и длинноволновых (ФС I) лучей красного света интенсивность фотосинтеза будет оптимальной.
Энергия, освобождающаяся при движении электронов от П680 до П700, используется для синтеза АТФ из AДФ и неорганического фосфата (фотофосфорилирование).
Наряду с нециклическим в мембранах хлоропластов функционирует циклический транспорт электронов, включающий в себя только ФС I и комплекс цитохромов Ь6 — f. В этом случае возбужденные молекулы П700 последовательно отдают электроны на А1, А2, АB, Фд, PQ, цитохром b6, FeSR, цитохром f, Пц и, наконец, П700. При этом НAДФ+ не восстанавливается. Освобождающаяся энергия используется для фосфорилирования AДP.
Интересно отметить, что комплекс цитохромов Ь6 — f функционирует в системе транспорта электронов при фотосинтезе аналогично комплексу III (цит. Ь, C1) в митохондриях.
Разница в уровнях энергии между П680 и П700 (> 50 кДж) вполне достаточна для фосфорилирования AДФ, так как величина высокоэнергетической фосфатной связи АТФ равна 30,6 кДж/моль (7,3 ккал). Перепады энергии в ФС I еще значительнее. Однако само по себе освобождение энергии при нециклическом и циклическом транспорте электронов не объясняет, каким образом осуществляется фотофосфорилирование.