Фотосинтез как основа энергетики биосферы
АТР участвует в фосфорилировании рибулозо-5-фосфата и ФГК, а также в реакциях, связанных с синтезом сахарозы и крахмала. NADPH необходим для восстановления ФГК до ФГА и для образования малата из щавелевоуксусной кислоты. Mg2+ нужен для функциональной активности РДФ-карбоксилазы, он также принимает участие в синтезе хлорофилла и белков.
Процесс фиксации С02 регулируется прежде всего светом.
Он активирует ряд ферментов цикла Кальвина: РДФ-карбоксилазу, дегидрогеназу 1,3-дифосфоглицериновой кислоты, фруктозо-1,6-дифосфатфосфатазу, седогептулозо-1,7-дифосфатфосфатазу и рибулозо-5-фосфаткиназу. Наиболее многообразна регуляция функционирования ключевого фермента цикла Кальвина — РДФ-карбоксилазы.
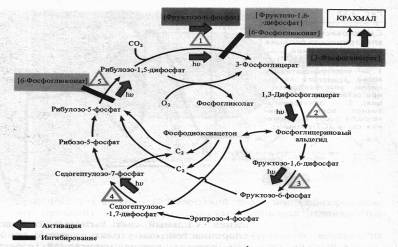
Помимо света, фермент активируется фруктозо-6-фосфатом, а ингибируется 6-фосфоглюконатом и фруктозо-1,6-дифосфатом. 6-Фосфоглюко-нат подавляет также активность завершающего цикл фермента — рибулозо-5-фосфаткиназы. Наконец, продукт цикла фиксации С02 — 3-фосфоглицериновая кислота положительно влияет на синтез крахмала. Повышение концентрации 02 в строме может привести к снижению интенсивности фиксации С02 за счет усиления фотодыхания.
Функциональные изменения в клетках мезофилла. Хлоропласты большинства растений способны перемещаться в клетке в зависимости от интенсивности и направления освещения. Сильный свет вызывает отрицательный фототаксис хлоропластов: они уходят от света, концентрируясь на боковых стенках клеток палисадной паренхимы; слабый свет вызывает положительный фототаксис. Предполагают, что освещенный хлоропласт меняет свое положение в клетке с помощью сократительных белков, связанных с оболочкой хлоропласта и взаимодействующих с сократительными белками цитоплазмы.
Мощное воздействие на клетку, определяющее ее метаболизм, оказывают образующиеся в хлоропластах ATP, NADPH и ассимиляты. При освещении отношение ATP/ADP в хлоропластах резко возрастает и вследствие быстрого обмена энергетическими эквивалентами почти тотчас же может воспроизводиться в цитоплазме, а через нее и в митохондриях. Так как оболочка хлоропласта лишь в слабой степени проницаема для АТР, перенос в цитоплазму высокоэнергетической фосфатной связи АТР осуществляется с помощью челночного механизма в результате реакции восстановления-окисления между ФГК и ФДА, так как скорость выхода триозофосфатов через мембраны оболочки хлоропласта приближается к скорости свободной диффузии. Одновременно это приводит к транспорту восстановительных эквивалентов (NADH).
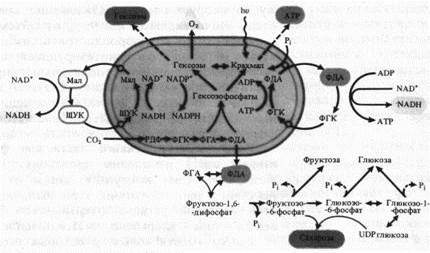
Переносчиком высокого восстановительного потенциала NADPH может служить система ЩУК-малат, каждый компонент которой легко проходит через мембраны, совершая циклические перемещения между хлоропластом и цитоплазмой. В пользу этой гипотезы говорит то, что в хлоропластах находятся NAD + - и NADP + -зависимые малатдегидрогеназы.
Появление АТР и восстановительных эквивалентов в цитоплазме приводит к активации на свету таких энергопотребляющих процессов, как синтез жирных кислот, восстановление нитратов и сульфатов. Общеклеточные нужды могут даже «отзывать» АТР с пути восстановления С02, конкурируя с реакциями цикла Кальвина.
Уже в первые секунды фотосинтеза происходит выход ассимилятов в цитоплазму. После кратковременного фотосинтеза с 14С02 в цитоплазме прежде всего появляются меченые фосфотриозы — ФГК и ФДА, а затем [14С] фруктозо-1,6-дифосфат. На изолированных хлоропластах показано, что оболочка хлоропласта практически непроницаема для сахарозы. По-видимому, основным местом синтеза сахарозы в клетке является не хлоропласт, а цитоплазма. На это указывает также присутствие основного сахарозосинтезирующего фермента сахарозофосфатсинтетазы и UDP-глюкозопирофосфорилазы преимущественно в цитоплазме.
Выход предшественников углеводов из хлоропластов в цитоплазму совершается в виде ФГК, ФГА и ФДА. Дальнейшие превращения, ведущие к синтезу гексозофосфатов и сахарозы, происходят уже в цитоплазме. Аналогичным превращениям могут подвергаться триозофосфаты и в хлоропластах, вследствие чего при достаточно интенсивном фотосинтезе в хлоропластах накапливается крахмал, служащий резервом углеводов, предназначенных для экспорта в цитоплазму во вторую очередь. Отложения крахмала часто бывают очень значительными (до 90% от объема хлоропласта).
Выходящие из хлоропластов ассимиляты могут использоваться в самой фотосинтезирующей клетке двумя путями: 1) в окислительно-восстановительных процессах, что обычно заканчивается освобождением С02, 2) для увеличения массы клетки в процессе ее роста и отложения запасных и вторичных веществ. Большая часть ассимилятов транспортируется из фотосинтезирующих клеток листа в другие органы и ткани.
Взаимодействие тканей листа при фотосинтезе. Под действием света изменения происходят не только в клетках мезофилла. Свет выступает одним из важнейших факторов регуляции работы устьиц. При включении света у большинства растений устьица открываются более широко, а при выключении — закрываются. Исключение из правила — растения семейства толстянковых, у которых ночью устьица открыты, а днем закрыты. На движения устьиц влияет и концентрация С02: ширина устьичных щелей увеличивается при снижении содержания С02 в межклетниках. Открывание устьиц под действием света связано с работой фотосинтетического аппарата в замыкающих клетках. У этиолированных растений свет не влияет на движения устьиц. В замыкающих клетках на свету включается механизм Н +-насоса в плазмалемме, усиливается поглощение К+ и синтез малата, в результате чего возрастает внутриклеточное осмотическое давление и устьица открываются.
Увеличение степени открывания устьиц приводит к усилению транспирации и, следовательно, к более интенсивному поступлению воды и веществ по ксилеме из корней в листья, что необходимо для нормальной функциональной активности клеток мезофилла.
Фотосинтетическая деятельность клеток мезофилла обогащает ткани листа сахарами и другими продуктами фотосинтеза. В результате возрастает функциональная активность проводящих пучков. Теоретически существуют два способа транспорта ассимилятов к проводящим пучкам: по симпласту (через плазмодесмы и цитоплазму последовательного ряда клеток) и по апопласту (по клеточным стенкам). Однако у многих видов растений между клетками мезофилла и флоэмы плазмодесмы развиты крайне слабо или совсем отсутствуют. В апопласте листовой пластинки может находиться около 1/5 сахаров, содержащихся в листе, и значительная доля свободных аминокислот. Клетки листовой паренхимы сравнительно легко выделяют ассимиляты в наружную среду и относительно слабо их поглощают. Клетки флоэмных окончаний, напротив, способны усиленно поглощать из внешних растворов сахара и аминокислоты против концентрационного градиента с помощью энергозависимых переносчиков. Есть основание считать, что, выходя из паренхимных клеток листа в клеточные стенки, сахароза расщепляется находящейся там инвертазой на гексозы (фруктозу и глюкозу), которые в проводящих пучках вновь образуют сахарозу.