Страница
7
Важное значение каротиноидов как дополнительных пигментов, поглощающих свет в синефиолетовой и синей частях спектра, становится очевидным при рассмотрении распределения энергии в спектре суммарной солнечной радиации на поверхности Земли.
Максимум радиации приходится на сине-голубую и зеленую части спектра (480 — 530 нм). В естественных условиях доходящая до поверхности Земли суммарная радиация слагается из потока прямой солнечной радиации на горизонтальную поверхность и рассеянной радиации неба. Рассеивание света в атмосфере происходит благодаря аэрозольным частицам (капли воды, пылинки и т. д.) и флуктуациям плотности воздуха (молекулярное рассеяние). Спектральный состав суммарной радиации в области 350 — 800 нм при безоблачном небе в течение дня почти не меняется. Объясняется это тем, что увеличение доли красных лучей в прямой солнечной радиации при низком стоянии Солнца сопровождается увеличением доли рассеянного света, в котором много сине-фиолетовых лучей. Атмосфера Земли в значительно большей степени рассеивает лучи коротковолновой части спектра (интенсивность рассеяния обратно пропорциональна длине волны в четвертой степени), поэтому небо выглядит голубым. При отсутствии прямого солнечного света (пасмурная погода) увеличивается доля сине-фиолетовых лучей. Эти данные указывают на важность коротковолновой части спектра при использовании наземными растениями рассеянного света и возможность участия каротиноидов в фотосинтезе в качестве дополнительных пигментов. В модельных опытах показана высокая эффективность переноса энергии света от каротиноидов к хлорофиллу а, причем этой способностью обладают молекулы каротинов, но не ксантофиллов.
Вторая функция каротиноидов — защитная. Впервые данные о том, что каротиноиды могут защищать молекулы хлорофилла от разрушения, были получены Д. И. Ивановским (1913). В его опытах пробирки, содержащие одинаковый объем раствора хлорофилла и разные концентрации каротиноидов, выставлялись на 3 ч на прямой солнечный свет. Оказалось, что чем больше каротиноидов было в пробирке, тем в меньшей степени разрушался хлорофилл. В дальнейшем эти данные получили многочисленные подтверждения. Так, бескаротиноидные мутанты хламидомонады на свету в атмосфере кислорода погибают, а в темноте при гетеротрофном способе питания нормально развиваются и размножаются. У мутанта кукурузы, у которого отсутствовал синтез каротиноидов, образующийся хлорофилл в аэробных условиях при сильном освещении быстро разрушался. В отсутствие кислорода хлорофилл не разрушался.
В настоящее время показано, что каротиноиды способны реагировать с хлорофиллом, находящимся в триплетном состоянии, предотвращая его необратимое окисление. При этом энергия триплетного возбужденного состояния хлорофилла превращается в теплоту:
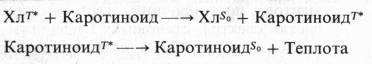
Кроме этого каротиноиды, взаимодействуя с возбужденным (синглетным) кислородом, который неспецифически окисляет многие органические вещества, могут переводить его в основное состояние:
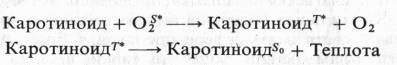
Менее ясна роль каротиноидов в кислородном обмене при фотосинтезе. У высших растений, мхов, зеленых и бурых водорослей осуществляется светозависимое обратимое дезэпоксидирование ксантофиллов. Примером такого превращения может служить виолаксантиновый цикл:
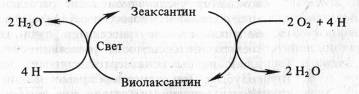
Значение виолаксантинового цикла остается невыясненным. Возможно, он служит для устранения излишков кислорода. Каротиноиды у растений выполняют и другие функции, не связанные с фотосинтезом. В светочувствительных «глазках» одноклеточных жгутиковых и в верхушках побегов высших растений каротиноиды, контрастируя свет, способствуют определению его направления. Это необходимо для фототаксисов у жгутиковых и фототропизмов у высших растений.
Каротиноиды обусловливают цвет лепестков и плодов у некоторых растений Производные каротиноидов — витамин А, ксантоксин, действующий подобно АБК, и другие биологически активные соединения. Хромопротеин родопсин, обнаруженный у некоторых галофильных бактерий, поглощая свет, функционирует в качестве Н+-помпы. Хромофорной группой бактериородопсина является ретиналь — альдегидная форма витамина А. Бактериородопсин аналогичен родопсину зрительных анализаторов животных.
4.3 Фикобилины. Их структура, функции и физиологическая роль
Синезеленые водоросли (цианобактерии), красные морские водоросли и некоторые морские криптомонады помимо хлорофилла а и каротиноидов содержат пигменты фикобилины. Наиболее известные представители фикобилинов — фикоэритробилины и фикоцианобилины. Первые преобладают у красных водорослей и определяют их цвет, вторые — у синезеленых.
Структура и свойства фикобилинов. По структуре фикобилины. (от греч. «phycos» — водоросль и лат. bilis — желчь) относятся к группе желчных пигментов — билинов (у животных представитель этой группы — билирубин). Это тетрапирролы с открытой цепью, имеющие систему конъюгированных двойных и одинарных связей. В своем составе они не содержат атомов магния или других металлов, а также фитола.
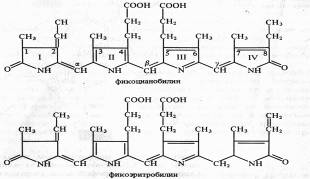
У фикоцианобилина пиррольные кольца соединены между собой метиновыми мостиками. I и IV пирролы имеют по одной карбонильной группе. Пиррольные кольца содержат следующие боковые радикалы: четыре метильных (у C1,3,6,7), винильную (у С2), этильную (у С8) и два остатка пропионовой кислоты (у С4 и С5).
Фикобилины являются хромофорными группами фикобилипротеинов — глобулиновых белков, с которыми в отличие от хлорофиллов они связаны прочными ковалентными связями. Фикобилипротеины делятся на три основные группы; 1) фикоэритрины — белки красного цвета с максимумом поглощения от 498 до 568 нм, 2) фикоцианины — сине-голубые белки с максимумами поглощения от 585 до 630 нм, 3) аллофикоцианины — синие белки с максимумами поглощения от 585 до 650 нм. Все эти хромопротеины обладают флуоресценцией с максимумами 575 — 578, 635 — 647 и 660 нм соответственно.
Фикобилипротеины водорастворимы, в клетках водорослей они локализованы в фшобилисомах — гранулах, расположенных на наружной поверхности фотосинтетических ламелл.
Значение фикобилинов. Максимумы поглощения света у фикобилинов находятся между двумя максимумами поглощения у хлорофилла: в оранжевой, желтой и зеленой частях спектра (см. рис. 4.3). Значение такого распределения максимумов поглощения становится понятным, если вспомнить оптические свойства воды, которая поглощает прежде всего длинноволновые лучи. На глубине 34 м в морях и океанах полностью исчезают красные лучи, на глубине 177 м — желтые, на глубине 322 м — зеленые и, наконец, на глубину свыше 500 м не проникают даже синие и фиолетовые лучи. В связи с таким изменением качественного состава света в верхних слоях морей и океанов обитают преимущественно зеленые водоросли, глубже — сине-зеленые и еще глубже — водоросли с красной окраской. В. Т. Энгельман назвал это явление хроматической комплементарной адаптацией водорослей. По его наблюдениям (1881 — 1884), наиболее интенсивная ассимиляция С02 у водорослей с различной окраской соответствует максимумам поглощения света пигментными системами этих водорослей. Русский исследователь Н. М. Гайдуков (1903) экспериментально показал, что если культуру синезеленой водоросли Oscillariasanctaвыращивать на свету разного спектрального состава, то у нее развивается дополнительная (комплементарная) окраска. При освещении зеленым светом водоросли становятся оранжево-красными, а при дейтвии красных лучей — зелеными. В настоящее время известно, что эти изменения цвета клеток связаны с изменениями в синтезе фикобилинов, принимающих участие в процессе фотосинтеза. Таким образом, у водорослей фикобилины — дополнительные пигменты, выполняющие вместо хлорофилла bфункции светособирающего комплекса. Около 90% энергии света, поглощенного фикобилинами, передается на хлорофиллы а. Явление хроматической комплементарной адаптации обнаружено далеко не у всех видов синезеленых и красных водорослей. У многих из них адаптация к изменяющемуся спектральному составу света обеспечивается изменением количества и состава хлорофиллов а.