Фотосинтез как основа энергетики биосферы
Флоэмные окончания выполняют в листе роль коллекторов, аккумулирующих ассимиляты против градиента концентрации. Ведущая роль в загрузке проводящих элементов ассимилятами принадлежит сопровождающим и паренхимным клеткам флоэмы, а у некоторых видов растений и более специализированным передаточным клеткам. Есть данные о том, что клетки флоэмных окончаний поглощают сахара с участием ионов Н +, которые выкачиваются из клеток благодаря работе энергозависимого Н +-насоса.
При перегрузке тканей листа ассимилятами интенсивность фотосинтеза снижается. Переполнение крахмалом пластид вызывает обратимые, а позднее и необратимые изменения структуры хлоропластов, что значительно ослабляет фотосинтез. Отрицательное действие избытка крахмала на фотосинтез в пластидах может быть объяснено: 1) механическим воздействием на тилакоиды, 2) ухудшением светового режима в хлоропласте, 3) сорбцией ферментов на зернах крахмала, 4) сорбцией ионов, особенно Mg2 + , необходимых для поддержания высокой активности ферментов, 5) ухудшением диффузии С02.
Регуляция процессов фотосинтеза в целом растении.
Реализация фотосинтетической функции в целом растении, с одной стороны, определяется значительной генетической и биохимической автономностью структур низших порядков (хлоропласт, клетка), а с другой — сложной системой интеграции и кооперативных связей фотосинтеза со всеми функциями растительного организма. Процессы онтогенеза обеспечивают постоянное существование в растительном организме так называемых аттрагирующих (притягивающих питательные вещества) зон. В аттрагирующих центрах происходит либо новообразование и рост структур, либо интенсивный однонаправленный синтез запасных веществ (плоды, клубни, луковицы). В обоих случаях состояние аттрагирующих центров определяет величину «запроса» на фотосинтез. Донор ассимилятов (фотосинтез) и их акцептор (процессы роста и отложения веществ в запас) представляют собой взаимосвязанную систему. Если внешние условия не лимитируют фотосинтез, то ведущая роль в его детерминации принадлежит именно эпигенетическим процессам (появлению и развитию новых органов). Это положение может быть проиллюстрировано многочисленными фактами. Так, удаление молодых початков у кукурузы, плодов у томатов, баклажанов или яблони вызывает значительное и устойчивое снижение фотосинтетической активности листьев. Удаление части листьев растения при сохранении прежней активности аттрагирующих центров обычно сопровождается увеличением фотосинтетической активности оставшихся листьев.
Механизм этих взаимоотношений основан на явлении метаболической репрессии фотосинтеза. Чем мощнее центры, аттрагирующие ассимиляты, тем эффективнее фотосинтезирующий лист освобождается от ассимилятов, что служит необходимым условием интенсивного фотосинтеза.
Во всех этих процессах важнейшую роль играют фитогормоны и эндогенные ингибиторы роста и метаболизма (некоторые полифенолы). Участие фитогормонов в процессах фотосинтеза можно представить в виде следующей схемы, предложенной А. Т. Мокроносовым (1983):
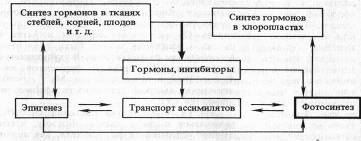
Согласно этой схеме, фитогормоны образуются в разных частях растений, в том числе в хлоропластах, и действуют на процессы фотосинтеза как дистанционно, так и непосредственно на уровне хлоропластов. Дистанционное действие осуществляется благодаря регулирующему влиянию фитогормонов на процессы роста и развития (эпигенез), на отложение веществ в запас, на транспорт ассимилятов, т. е. на формирование и активность аттрагирующих центров. С другой стороны фитогормоны оказывают прямое действие на функциональную активность хлоропластов через изменение состояния мембран, активность ферментов, генерацию трансмембранного потенциала. Доказана также роль фитогормонов, в частности цитокинина, в биогенезе хлоропластов, синтезе хлорофилла и ферментов цикла Кальвина.
Ряд фитогормонов (индолилуксусная кислота, гиббереллины, абсцизовая кислота), а также некоторые физиологически активные фенолы образуются в тканях листа. Цитокинин же, от которого во многом зависит формирование листа и его структур, поступает из других частей растения, прежде всего из корня. Такая система создает взаимозависимость всех органов, обеспечивая регуляцию функциональной активности в целом растении.
9 Зависимость процесса фотосинтеза от факторов внешней среды
При оценке действия внешних факторов необходимо различать два уровня. Первый их них генетический, который определяется влиянием факторов на генетический аппарат и экспрессию генов. Второй уровень обусловлен прямым действием внешних факторов на отдельные реакции фотосинтеза. Ответная реакция организма на изменение внешних факторов может быть быстрой, когда определяется непосредственно их воздействие на фотосинтетический аппарат, и более медленной, когда в новых условиях происходит формирование структур. Действие экзогенных факторов взаимосвязано и взаимообусловлено и реализуется через конкретные механизмы, сопряженные со всем комплексом физических, фотохимических и энзиматических реакций фотосинтеза. Познание этих механизмов необходимо для глубокого понимания основных закономерностей и оптимальных условий функционирования фотосинтетического аппарата как целостной системы.
Рассмотрим отдельно влияние на фотосинтез основных факторов внешней среды, хотя в природе они действуют на растение одновременно, и продуктивность растения является интегральной функцией совместного действия ряда экологических факторов.
Влияние интенсивности и спектрального состава света на фотосинтез
Интенсивность света и фотосинтез. Зависимость фотосинтеза о т л у ч и с т о й энергии является наиболее очевидной и существенной. Уже в ранних работах К.А.Тимирязева и других исследователей установлено отсутствие линейной зависимости между активностью процесса фотосинтеза и напряженностью действующего фактора. Зависимость активности фотосинтеза от интенсивности света — световая кривая фотосинтеза — имеет форму логарифмической кривой. Прямая зависимость скорости процесса от притока энергии имеет место только при низких интенсивностях света. В области насыщающих интенсивностей света дальнейшее увеличение освещенности не увеличивает скорость фотосинтеза.
Эти данные послужили основанием для представлений о включении в процесс фотосинтеза наряду с фотохимическими, световыми реакциями также темновых, энзиматических реакций, ограничивающее действие которых начинает проявляться особенно заметно при высоких, насьпдающих интенсивностях света. Результаты опытов А. А. Рихтера и Р. Эмерсона с прерывистым светом позволили оценить скорость световых и темновых реакций фотосинтеза: соответственно 10-5 и 10-2 с. Эти значения были полностью подтверждены в лаборатории Х.Витта (Witt, 1966) с использованием высокочувствительных методов импульсной спектрофотометрии.
При проведении физиологических исследований анализ световой кривой фотосинтеза дает информацию о характере работы фотохимических систем и ферментативного аппарата. Угол наклона кривой характеризует скорость фотохимических реакций: чем он больше, тем активнее система использует энергию света. По углу наклона линейного участка можно вести приближенные расчеты расхода квантов на восстановление моля С02. Скорость фотосинтеза в области насыщающей интенсивности света характеризует мощность систем поглощения и восстановления С02 и в значительной мере определяется концентрацией углекислоты в среде. Чем выше расположена кривая в области насыщающих интенсивностей света, тем более мощным аппаратом поглощения и восстановления углекислоты обладает система.