Фотосинтез как основа энергетики биосферы
Лист, как правило, имеет плоскую форму и дорсовентральное строение. Пластинчатая форма листа обеспечивает наибольшую поверхность на единицу объема ткани, что создает наилучшие условия для воздушного питания.
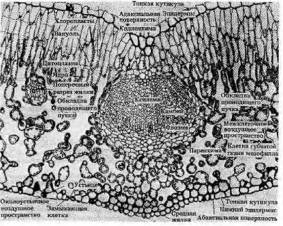
Поперечный разрез листовой пластинки и средней жилки листа Бирючины – типичного двудольного растения
Важнейшая ткань листа — мезофилл, где осуществляется фотосинтез. Покрывающий лист эпидермис, клетки которого, за исключением замыкающих клеток устьиц, не содержат хлоропластов, защищает ткани листа, регулирует газообмен и транспирацию. Система разветвленных проводящих пучков необходима для снабжения тканей листа водой, минеральными и некоторыми органическими веществами и для оттока ассимилятов в другие части растения. Мезофилл обычно дифференцирован на две ткани — палисадную (столбчатую), расположенную под верхним эпидермисом, и губчатую, находящуюся в нижней стороне листа. В палисадном мезофилле клетки вытянуты перпендикулярно поверхности листа и расположены в один или несколько слоев. Клетки губчатого мезофилла связаны друг с другом более рыхло из-за больших межклетников. У большинства растений устьица находятся нанижней стороне листа, обширные межклетники губчатой паренхимы обеспечивают газообмен. Благодаря развитой системе межклетников мезофилл обладает громадной поверхностью, во много раз превышающей наружную поверхность листа.
Палисадная ткань, обращенная к свету, содержит большую часть всех хлоропластов листа и выполняет основную работу по ассимиляции С02. Число слоев клеток мезофилла в листе в большой степени зависит от интенсивности освещения. Листья, выросшие на ярком свету, имеют как правило, более развитый столбчатый мезофилл. У растений засушливых ареалов — ксерофитов — палисадная паренхима часто расположена по обеим сторонам листа, а губчатая сильно редуцирована или совсем отсутствует.
3.2 Состав, строение, функции, возникновение и развитие хлоропластов
Хлоропласты имеют зеленый цвет, обусловленный присутствием основного пигмента — хлорофилла. Хлоропласты содержат также вспомогательные пигменты — каротиноиды (оранжевого цвета). По форме хлоропласты — это овальные линзовидные тельца размером (5—10) * (2—4) мкм. В одной клетке листа может находиться 15—20 и более хлоропластов, а у некоторых водорослей — лишь 1-2 гигантских хлоропласта (хроматофора) различной формы.
Хлоропласты ограничены двумя мембранами — наружной и внутренней. Наружная мембрана отграничивает жидкую внутреннюю гомогенную среду хлоропласта — строму (матрикс). В строме содержатся белки, липиды, ДНК (кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные и белковые зерна) а также ферменты, участвующие в фиксации углекислого газа.
Внутренняя мембрана хлоропласта образует впячивания внутрь стромы — тилакоиды, или ламеллы, которые имеют форму уплощенных мешочков (цистерн). Несколько таких тилакоидов, лежащих друг над другом, образуют грану, и в этом случае они называются тилакоидами граны. Именно в мембранах тилакоидов локализованы светочувствительные пигменты, а также переносчики электронов и протонов, которые участвуют в поглощении и преобразовании энергии света.
Хлоропласты в клетке осуществляют процесс фотосинтеза.
Лейкопласты — мелкие бесцветные пластиды различной формы. Они бывают шаровидными, эллипсоидными, гантелевидными, чашевидными и т. д. По сравнению с хлоропластами у них слабо развита внутренняя мембранная система. Лейкопласты в основном встречаются в клетках органов, скрытых от солнечного света (корней, корневищ, клубней, семян). Они осуществляют вторичный синтез и накопление запасных питательных веществ — крахмала, реже жиров и белков.
Хромопласты отличаются от других пластид своеобразной формой (дисковидной, зубчатой, серповидной, треугольной, ромбической и др.) и окраской (оранжевые, желтые, красные). Хромопласты лишены хлорофилла и поэтому не способны к фотосинтезу. Внутренняя мембранная структура их слабо выражена.
Хромопласты присутствуют в клетках лепестков многих растений (лютиков, калужниц, нарциссов, одуванчиков и др.), зрелых плодов (томаты, рябина, ландыш, шиповник) и корнеплодов (морковь, свекла), а также листьев в осеннюю пору. Яркий цвет этих органов обусловлен различными пигментами, относящимися к группе каргиноидов, которые сосредоточены в хромопластах.
4 Пигменты хлоропластов
Пигменты — важнейший компонент аппарата фотосинтеза. Изучение растительных пигментов резко ускорилось благодаря работам русского физиолога растений М. С. Цвета. Пытаясь найти способ разделения пигментов на индивидуальные вещества, Цвет в 1901 — 1903 гг. открыл принципиально новый метод, который он назвал адсорбционной хроматографией. Через колонку с сорбентом пропускается растворитель с растворенными веществами. Так как вещества различаются по степени адсорбции, они перемещаются по колонке с разной скоростью. В результате происходит разделение веществ. Этот прием широко используется в современной биохимии, химии и в некоторых отраслях промышленности. С помощью хроматографического метода Цвет обнаружил два хлорофилла — а и bи разделил желтые пигменты листа на три фракции.
Пигменты пластид относятся к трем классам веществ: хлорофиллам, фикобилинам и каротиноидам.
4.1 Хлорофиллы: состав, структура, биосинтез, оптические и химические свойства, значение
Впервые хлорофилл в кристаллическом виде был описан русским физиологом и ботаником И. П. Бородиным в 1883 г. В дальнейшем оказалось, что это не сам хлорофилл, а несколько видоизмененная его форма — этилхлорофиллид.
Польские биохимики М. Ненцкий и Л. Мархлевский (1897) обнаружили, что основу молекулы хлорофилла, как и гема гемоглобина, составляет порфириновое кольцо. Таким образом было показано принципиальное структурное сходство этих пигментов у растений и животных.
Немецкий химик Р. Вильштеттер в 1906—1914 гг. установил элементарный состав хлорофилла а — C55H72O5N4Mg и хлорофилла b — C55H70O6N4Mg, а немецкий биохимик Г. Фишер в 1930—1940 гг. полностью расшифровал структурную формулу хлорофилла. В 1960 г. химики-органики Р.Б. Вудворд (США) и М. Штрель (ФРГ) осуществили искусственный синтез хлорофилла. Хлорофилл — сложный эфир дикарбоновой кислоты хлорофиллина, у которой одна карбоксильная группа этерифицирована остатком метилового спирта, а другая остатком одноатомного непредельного спирта фитола. Структурная формула хлорофилла а представлена на рисунке:
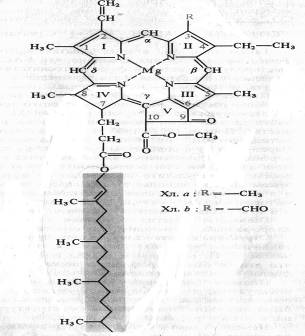
Четыре пиррольных кольца (I — IV) соединены между собой метановыми мостиками, образуя порфириновое ядро. Наружные атомы углерода в тетрапиррольном цикле обозначены цифрами 1 — 10. Атомы азота пиррольных колец четырьмя координационными связями взаимодействуют с атомом магния. В структуре порфиринового ядра есть также циклопентановое кольцо (V), образованное остатком кетопропионовой кислоты и содержащее химически активную карбонильную группу у С9 и метилированную карбоксильную группу у С10. Структура, состоящая из тетрапиррольного и циклопентанового колец, получила название форбина. Боковая цепь IV пиррольного кольца включает в себя пропионовую кислоту, связанную сложноэфирной связью с полиизопреновым непредельным спиртом фитолом (С20Н39ОН). У 1, 3, 5 и 8-го углеродов пиррольных колец имеются метальные группы, у 2-го — винильная, у 4-го — этильная группа. Порфириновое кольцо представляет собой систему из девяти пар конъюгированных (сопряженных) чередующихся двойных и одинарных связей с 18 делокализованными p-электронами. Хлорофилл b отличается от хлорофилла а тем, что у 3-го углерода вместо метильной находится формильная (—СНО) группа. Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами в молекуле хлорофиллов образуются соответствующие феофетины.