Свет и его роль в жизни растений и животных
Метаболическая компартментализация аминокислот, в частности аминокислот глутаминовой группы, особенно ярко проявляется в субклеточной локализации ферментов в ГАМК-ергическом нейроне и в астроцитах. Так, пируваткар-боксилаза локализована преимущественно в астроцитах, в то время как пируватдегидрогеназный комплекс более активен в нейронах, чем в астроцитах. Преимущественная локализация пируватдегидрогеназы в нейронах ответственна в конечном счете за низкое включение углерода глюкозы, лактата и глицерина в глутамин. Глутамин-синтетаза преимущественно локализована в астроцитах, а глутаминаза - в нейронах. Глутамат, поглощаясь астроцитами, превращается в глутамин в глутамин-синтетазной реакции. Глутамин, выйдя из астроцитов и входя в нейроны, образует глутамат и далее ГАМК. Таким образом, глутамин и ГАМК осуществляют коммуникацию между большим и малым метаболическими компартментами.
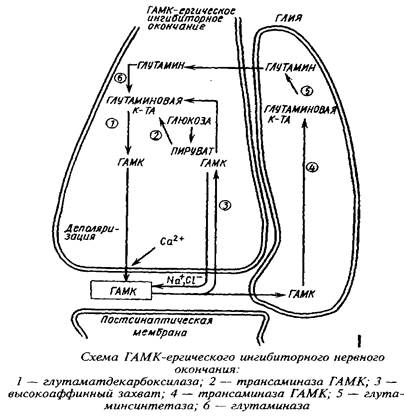
Помимо нейронально-глиального транспорта аминокислот в последние годы установлена возможность перемещения свободных аминокислот от проксимального к дистальному концу нейрона. Так, глутамат, введенный в мозг, передвигается вдоль аксонов двигательных нейронов и оказывается в мышечных нервных окончаниях.
7. Глицин и пути его обмена
Глицин участвует не только в биосинтезе белков, но и в других многочисленных биосинтетических процессах, таких, как образование пуринов, порфиринов, креатина, этаноламина, холина, глутатиона и др. Глицин функционирует также в качестве ингибиторного трансмиттера главным образом в спинном мозге.
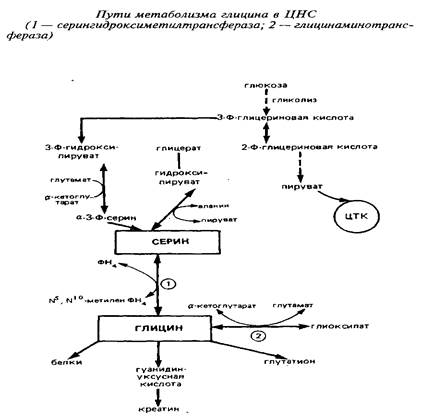
Так как потребление глицина в нервной ткани относительно велико, а поступление его из крови происходит медленно, значительная часть глицина синтезируется в мозге de novo. Глюкоза и серии являются главными источниками глицина в ЦНС. Серии может образовываться из глюкозы через 3-фосфоглице-риновую кислоту. Кроме того, серии сравнительно быстро поступает из циркулирующей крови. Синтез глицина de novo происходит в нервной ткани из серина путем обратимой N, W-метилентетрагидрофолат-тет-рагидрофолатзависимой трансформации при участии фермента серингидроксил1етилтрансферазы. Реакция катализируется серингидроксиметилтрансферазой и протекает следующим образом:
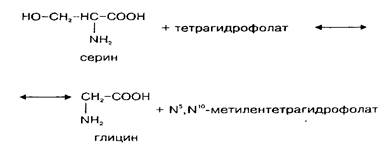
Этот фермент относится к пиридоксальзависимым при оптимальной активности в нем содержится 6 молекул пиридоксальфосфата. Активность фермента в метаболических пулах головного мозга относительно постоянна, высокая активность его обнаружена в спинном мозге и в мозжечке. Активность в сером веществе спинного мозга больше, чем в белом, причем в вентральном сером веществе она значительно выше, чем в дорзальном. Это коррелирует с содержанием глицина.
Другим источником синтеза глицина в нервной системе является глиоксиловая кислота, однако вклад ее в синтез глицина в головном мозге in vivo не может быть значительным, так как ее уровень в мозге низок.
В нервной ткани существует по крайней мере три пути катаболизма глицина. Первый состоит в том, что реакция превращения серина в глицин легко обратима в ткани мозга и серин-гидроксиметилтрансфераза может выступать в качестве энзима деградации глицина. Кроме того, в ЦНС представлены оксидазы аминокислот, которые могут использовать в качестве субстрата наряду с другими аминокислотами глицин:
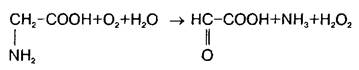
Третья система распада глицина локализована исключительно в митохондриях и является нетипичной декарбоксилазой аминокислот, так как зависит и от НАД+, и от тетрагидрофолата. Расщепление глицина на одноуглеродные фрагменты протекает по схеме:
![]()
Важно отметить образование в этих реакциях метилентетра-гидрофолата, который может быть использован в мозге как источник одноуглеродных фрагментов. То же следует подчеркнуть применительно к описанной серингидроксиметилтрансферазной реакции.
При участии глицин-расщепляющей системы глицин распадается на метилентетрагидрофолат, диоксид углерода и аммиак, затем происходит окисление метилен-ТГФ с образованием СОз - окончательного продукта распада глицина.
Как уже упоминалось, глицин является ингибиторным трансмиттером в спинном мозге. В других районах депрессорное действие глицина проявляется слабо. Поэтому спинной мозг имеет высокоаффинную и низкоаффинную систему захвата глицина, в то время как кора головного мозга содержит только низкоаффинную систему.
Интересно отметить, что повышенный уровень глинина обнаружен в эпилептогенных районах мозга человека, удаленных хирургическим путем. Он накапливается также в эпилептогенных районах мозга у животных с вызванными кобальтом припадками, причем тяжесть припадков пропорциональна накоплению глицина. Возможно, это - компенсаторные процессы.
Высокий уровень глицина в плазме крови или в моче обычно свидетельствует о нарушении мозговых функций. Гиперглицинемия развивается в раннем возрасте и сопровождается эпизодическими рвотами, подавлением двигательной активности, нарушением ЭЭГ и часто кончается смертью. Известны два типа гиперглииинемии - кетотическая и некетотическая, которая в большинстве случаев тоже летальна. Кетотическая гиперглицинемия сопровождается губчатой дегенерацией белого вещества мозга и задержкой миелинизации.
8. Серосодержащие аминокислоты
Метионин представляет особый интерес как источник метильных групп. Полученный с пищей, а также образованный в других тканях, метионин поступает в мозг через систему активного транспорта больших нейтральных аминокислот. Концентрация метионина в целом мозге сравнительно низка - от 10 до 100 нмоль/г сырой массы у различных видов животных. Региональные различия в концентрации метионина невелики. Влияние диеты на концентрацию метионина в мозге также незначительно из-за конкурентных отношений с нейтральными аминокислотами за транспортные системы. Метионин в пуле свободных аминокислот утилизируется на 80% для синтеза белка.
Метаболизм свободного метионина до цистеина начинается с образования S-аденозилметионина, реакция катализируется метионин-аденозилтрансферазой. S-Аденозилметионин является главным донором метальных групп в мозге, необходимых для метилирования катехоламинов, гистамина, фосфатидилэтаноламина, нуклеиновых кислот.
Процессам метилирования отводится важная роль: в проведении сигнала через мембрану, в регулировании жидкостно-сти мембраны и, наконец, в процессах метилирования ДНК. Последние считают вероятными участниками механизмов долговременной памяти. В то время как первая половина цикла превращения метионина связана главным образом с метилированием, вторая часть его ассоциирована в основном с нейротрансмиттерной и нейромо-дуляторной функцией. Оказалось, что 20% серусодержащих аминокислот локализовано в синаптосомах.
