Свет и его роль в жизни растений и животных
Постоянство суммарного аминокислотного пула головного мозга сопровождается региональной неоднородностью их содержания, что отражает морфологическую, физиологическую и функциональную гетерогенность этого органа. Наиболее неравномерно распределены аминокислоты, выполняющие функцию нейротрансмиттеров, такие, как глутаминовая кислота, таурин, ГАМК, глицин и др.
Таблица 2
Содержание аминокислот в различных областях мозга кошки
|
Аминокислоты |
Тал a wye |
Средний мозг |
Мозолистое тело |
Кора височной доли |
Мозжечок |
|
Глутаминовая |
12,36 |
9,71 |
10,58 |
12,93 |
12,63 |
|
Аспарагиновая |
2,71 |
4,06 |
1,41 |
3,09 |
2,85 |
|
Таурин |
1,06 |
1,62 |
2,99 |
1,89 |
3,12 |
|
Глицин |
1,72 |
2,77 |
0,614 |
1,25 |
1,49 |
|
Алании |
0,591 |
1,09 |
0,704 |
0,863 |
0,895 |
|
ГАМК |
3,65 |
5,81 |
0,961 |
1,39 |
1,49 |
|
Тирозин |
0,05 |
0,059 |
0,049 |
0,039 |
0,06 |
|
Валин |
0,145 |
0,152 |
0,096 |
0,117 |
0,097 |
|
Лизин |
0,278 |
0,379 |
0,268 |
0, 194 |
0,219 |
Различные органеллы клеток головного мозга контролируют уровень аминокислот, накапливая их часто против концентрационных градиентов.
Постоянство качественного и количественного состава аминокислот в метаболических фондах мозга обеспечивается такими взаимосвязанными процессами, как поступление аминокислот из циркулирующей крови, отток их из мозга в кровь и участие в реакциях внутриклеточного метаболизма. В организме все эти процессы сбалансированы слаженным функционированием гомеостатических механизмов, гематоэнцефалического барьера и мембранным транспортом.
Транспорт аминокислот в мозг - многоступенчатый процесс. Прежде всего происходит транспорт через гематоэнцефалический барьер, локализованный в эндотелии мозговых капилляров, затем осуществляется транспорт из внеклеточной жидкости в клетки мозга, а далее - в субклеточные органеллы. Существуют системы активного транспорта аминокислот не только в мозг, но и из него, - обе они энергозависимы.
Исследование конкурентных отношений в транспорте аминокислот выявило наличие восьми классов транспортных систем, которые существуют для аминокислот с родственной структурой и зависят от ионного заряда и размеров их молекул. В ряде случаев одна аминокислота может транспортироваться с участием нескольких транспортных систем, выбор той или иной системы определяется составом аминокислотного пула. Для мембранного транспорта аминокислот характерен ряд особенностей: а) перенос аминокислот часто происходит против высоких концентрационных градиентов; б) этот процесс энергозависим; в) на него влияют температура и рН среды; г) он ингибируется анаэробиозом и ферментными ядами; д) перенос аминокислот связан с активным мембранным транспортом ионов, в частности, он Na-зависим; е) обнаружено конкурентное торможение мембранного транспорта одних аминокислот другими и др. Такие конкурентные взаимодействия играют важную роль в патологии, когда изменяется уровень индивидуальных аминокислот в крови. Ниже мы приведем примеры таких патологических состояний.
Уровень специфичности транспортных систем для разных аминокислот неодинаков. Особенно велика специфичность и мощность систем для аминокислот, выполняющих роль нейротрансмиттеров. Эти системы не только обеспечивают пластические и энергетические нужды клетки, но служат такие для специфического процесса быстрого снижения концентрации нейротрансмиттера в зоне синоптической щели. Высокоизбирательное поглощение нейротрансмиттера осуществляется как пресинаптической областью, так и клетками окружающей глии.
Еще один своеобразный механизм транспорта аминокислот связан с метаболизмом широко распространенного во всех тканях, в том числе и в нервной, трипептида глутатиона, цикл синтеза и деградации которого известен под названием у-глутамильного цикла. Наиболее интересным и ключевым ферментом этого цикла является у-глуталшлтранспептидаза, прочно связанная с клеточной мембраной. Этот энзим способен переносить у-глутамильную группу глутатиона, находящегося внутри клетки, на аминокислоту, локализованную с наружной стороны мембраны, и переносить образующийся дипептид внутрь клетки. Следующий фермент этого цикла - у-глутамилциклотрансфераза высвобождает аминокислоту. Таким образом, у-глутамил транспептидазная реакция является одним из механизмов транспорта аминокислот внутрь клетки.
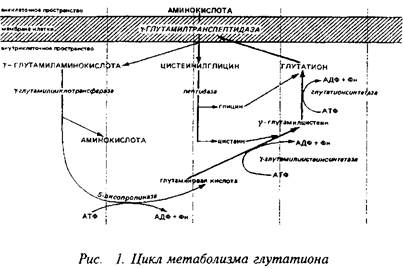
При нормальных условиях скорость транспорта аминокислот не лимитирует непосредственно их метаболизм, так как скорости синтеза и деградации ниже скорости транспорта. Поэтому аминокислоты и аккумулируются мозгом, формируя пул свободных аминокислот. Без пополнения извне пул свободных аминокислот довольно быстро истощается. Так, количество аминокислот, которое используется для синтеза белков мозга, нейропептидов и нейромедиаторов в течение 30 мин, равно общему церебральному пулу большинства свободных аминокислот.