Свет и его роль в жизни растений и животных
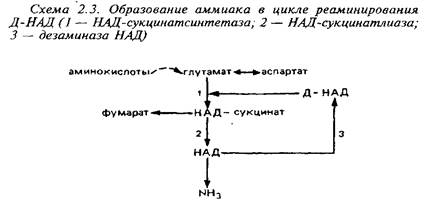
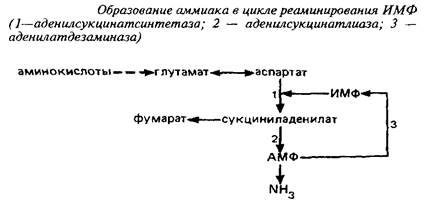
Вне митохондрий действует другой циклический процесс образования аммиака, в котором аспартат реаминирует инозинмонофосфат.
Для удаления аммиака в ЦНС служит глутаминсинтетазная реакция.
Глутаминсинтетаза катализирует реакцию:
![]()
Этот энзим в мозге животных находится в более высокой концентрации, чем в других органах, составляя 0,2% от общего белка мозга. Энзим требует АТФ и Mg+ и подавляется глицином и аланином. Км для аммония - порядка 0,39 мМ, т.е. при нормальной концентрации аммония в мозге фермент работает в режиме полунасыщения. В нормальных физиологических условиях, когда имеется достаточный уровень АТФ, глу-таминсинтетазная реакция направлена в сторону связывания аммиака.
Образование глутамина является важным механизмом детоксикации аммония, к которому мозг чрезвычайно чувствителен и накопление которого губительно для ЦНС. В частности, повышение аммиака в мозге до концентрации 0,6 мМ сопровождается судорогами. Системное введение солей аммония вызывает конвульсии и увеличение содержания глутамина в мозге. В случае серьезных повреждений печени повышается концентрация аммония и глутамина в спинномозговой жидкости - в этих случаях наблюдается кома. Симптомы печеночной комы смягчаются введением глутамата. Основная часть глутаминсинтетазы локализована в глиальных клетках и лишь небольшая часть ее представлена в нервных окончаниях.
Дезаминирование глутамина катализируется глутаминазой, ферментом, наиболее активным в нейронах, где он локализован в митохондриях. Следует отметить, что активность этого фермента в головном мозге невелика; продукты реакции - глутаминовая кислота и аммоний - тормозят активность фермента.
Предполагается участие этого фермента в мембранном транспорте глутамата. Известно, что биологические мембраны более проницаемы для глутамина, чем для глутамата, и глутаминаза может участвовать в превращении глутамина крови во внутриклеточный глутамат. Глутаминаза играет важную роль также в регуляции содержания глутамата в нервных окончаниях. Тот факт, что глутаминсинтетаза локализована в основном в глиальных клетках, а глутаминаза наиболее активна в нейронах, а также то, что глутамин оказался главным предшественником глутамата и ГАМ К, выполняющих трансмиттерную функцию, послужил основанием для концепции о существовании глушаминового цикла, Глутамат, поглощаясь глиальными клетками, превращается в глутамин в синтетазной реакции, последний входит в нейроны, образуя там глутаминовую кислоту. Таким образом, глутамин служит глиально-нейронааьным транспортером глутамата.
Другой важной функцией глутамата является его участие в синтезе белков и биологически активных пептидов. Глутамат и глутамин составляют вместе от 8 до 10% общих аминокислотных остатков в гидролизате белков мозга. В частности, два хорошо изученных мозгоспецифичных белка - S-100 и 14-3-2 - содержат особенно высокую долю глутаминовой кислоты. Глутамат является также составной частью ряда малых и средних регуляторных пептидов мозга. Это прежде всего глутатион и ряд у-глутамильньгх дипептидов. Некоторые нейропептиды содержат циклическое производное глутамата - аироглутамат в качестве N-терминального остатка, который предохраняет эти пептиды от протеолиза. К таким "пептидам относятся люлибе-рин, тиролиберин, нейротензин, бомбезин и др. .
Введение глутамата в различные районы мозга приводит либо к судорожной активности, либо к распространяющейся депрессии, даже если количество его мало по сравнению с нормальной концентрацией глутамата в мозге. Глутамин не вызывает такого эффекта. При внутривенном введении глутамат может вызвать гибель клеток в определенных районах ЦНС, особенно вокруг желудочков мозга, где менее развит гематоэнцефалический барьер. Нейроны незрелых животных, у которых еще отсутствует высокоразвитый гематоэнцефалический барьер, также очень чувствительны к глутамату. Оральное введение больших количеств глутамата не действует на ЦНС большинства людей, а соли глутамата широко используются в качестве пищевой приправы. Однако у некоторых лиц обнаруживается повышенная чувствительность к глутамату натрия, он вызывает сенсорные и моторные нарушения, включая ощущение жжения, напряжение лица, боль в грудной клетке и головную боль. Эти симптомы известны как "синдром китайских ресторанов", так как глутамат натрия широко используется в китайской кухне. Многие аналоги глутамата токсичны.
Остановимся на некоторых сторонах нейротрансмиттерной функции глутамата. Для того чтобы глутамат эффективно функционировал в качестве нейротрансмиттера, его модальная внеклеточная концентрация должна быть ниже той, которая вызывает деполяризацию мембран. В действительности она колеблется от 1 до 10 мкМ; такая низкая внеклеточная концентрация глутамата поддерживается активным транспортом в нейроны и особенно в глиальные клетки.
В процессе выхода глутамата в синаптическую щель концентрация его там значительно повышается - до 1 мМ.
Последующий обратный захват глутамата нейронами и астроцитами осуществляется с участием Na-зависимых высокоаффинных переносчиков, из синаптической щели глутамат удаляется в основном путем захвата астроцитами. Для функционирования глутамата в качестве нейротрансмиттера необходимо постоянное пополнение его пула в нервных окончаниях.
Предшественниками трансмиттерного пула глутамата могут быть глюкоза и а-кетоглутарат.
Глутамат может также образовываться из орнитина и аргинина. Но основным источником нейротрансмиттерного глутаматного пула, по данным изотопных исследований, оказался глутамин, который синтезируется в основном в астроцитах, где локализована глутаминсинтетаза.
Далее он легко транспортируется через мембрану астроцитов и с помощью активных переносчиков достигает нервных окончаний.
4. N-Ацетиласпарагиновая кислота
Одним из доминирующих компонентов пула свободных аминокислот мозга является N-ацетиласпарагиновая кислота

Ее концентрация у большинства видов животных в два раза превышает таковую аспарагиновой кислоты. В ненейрональной ткани обнаружены только следы АцА. Она находится в более высокой концентрации в сером веществе по сравнению с белим, представлена также в периферической нервной системе, в сетчатке. Ее концентрация низка при рождении и повышается в процессе развития животного.
АцА образуется с участием ацетил-КоА. Энзим, катализирующий эту реакцию, очищен и изучен. Точная функция АцА в мозге еще не ясна, хотя имеются предположения, что она является частью внутриклеточного фиксированного пула анионов или резервуаром ацетильных групп, а также источником N-ацетилированных конечных групп для синтеза определенных бел-
ков и пептидов мозга. Показано, что ацетильные группы экзогенной АцА кислоты служат предпочтительным источником углерода для синтеза жирных кислот в развивающемся мозге. В головном мозге оказалось два пространственно разобщенных фонда АцА: малый, высокоактивный, локализованный в глии, и большой, медленно обменивающийся, - в нейронах.