Клеточная биотехнология
Кроме изменения плоидности, культивирование клеток и тканей растений in vitro вызывает появление в клетках хромосомных аббераций. Последние сказываются на биологических особенностях культивируемых тканей, изменяя их внешний вид, обмен веществ, скорость роста. Наряду с видимыми под микроскопом хромосомными мутациями в культивируемых клетках могут возникать изменения, не выявляемые микроскопически. Эти изменения могут затрагивать как незначительные участки хромосом, так и структуру генов. Генные мутации выявляются по изменению морфологии и физиолого-биохимических свойств клеток.
Каковы же причины генетической нестабильности культивируемых клеток? Таких причин несколько. Прежде всего – это генетическая неоднородность исходного материала (гетерогенность экспланта). У многих растений дифференцированные ткани характеризуются наличием клеток разной плоидности и лишь активно пролиферирующие в течение онтогенеза ткани, такие, как верхушечные меристемы, камбий и другие, остаются всегда диплоидными. Другой причиной может быть длительное пассирование тканевых и клеточных культур, приводящее к накоплению в них генетических изменений, в том числе к неравномерному изменению плоидности. Нарушение коррелятивных связей при изолировании участков тканей растений и помещении их на питательную среду также приводит к генетической нестабильности клеток. Подобные результаты могут быть связаны и с влиянием на генетический аппарат клетки входящих в состав питательных сред фитогормонов. В качестве гормонов в питательные среды для каллусообразования обязательно входят ауксины и цитокинины. О мутагенном действии этих веществ известно из целого ряда работ. Наиболее активным мутагенным препаратом является 2,4-Д (2,4-Дихлорфеноксиуксусная кислота – синтетический аналог индолилуксусной кислоты), входящий в состав большинства питательных сред. Цитокинины, в частности кинетин, способствуют полиплоидизации клеток. Не исключено, что возникновение генетических аббераций вызвано накоплением вторичных метаболитов, и в частности полифенолов.
Существует несколько путей, по которым может идти развитие клетки после ее дедифференцировки.
Первый путь – это вторичная регенерация целого растения, возможна дифференцировка на уровне клеток, тканей, органов.
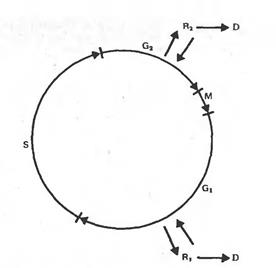
Второй путь - это утрата клеткой способности к вторичной дифференцировке и регенерации растения, стойкая дедифференцировка, приобретение способности расти на среде без гормонов, т.е. превращение в опухолевую. Такими свойствами часто характеризуются клетки старых пересадочных культур. На рисунке изображены фазы клеточного цикла, и показано, в каких из них клетки могут выйти из митотического цикла, и перейти в дифференцированное состояние и соответственно вернуться в цикл при дедифференцировке и индукции их к делению. Обычно клетки переходят к специализации из фазы G1.
Третий путь - это нормальный цикл развития каллусной клетки, заканчивающийся ее старением и отмиранием. В этом случае клетка претерпевает вторичную дифференцировку и прекращает делиться (стационарная фаза роста). Однако такая дифференцировка не ведет к морфогенезу, а закрепляет за ней свойства старой каллусной клетки. В культуре каллусных тканей морфогенезом называют возникновение организованных структур из неорганизованной массы клеток.
Существует два основных типа морфогенеза. В культуре тканей он может проявляться в виде органогенеза (образования монополярной структуры, т.е. отдельных органов); корневого, стеблевого, реже флорального (цветочного) или листового, в виде соматического эмбриогенеза (образования биполярных зародышеподобных структур из соматических клеток).
В случае органогенеза сначала регенерируют отдельные органы, а затем уже из них - целые растения, исключение составляет корневой органогенез. В результате соматического эмбриогенеза в отличие от органогенеза сразу образуется зародыш, имеющий как меристему корня, так и меристему верхушечной почки, из которого в дальнейшем развивается целое растение.
Согласно концепции тотипотентности, если мы получаем каллус из клеток лепестка цветка, или из клеток сердцевинной паренхимы стебля, или из клеток любой ткани, то в принципе каждая такая клетка может регенерировать целое растение. Однако свойство тотипотентности не всегда реализуется, так как потенциальные возможности клеток разных типов проявляются неодинаково. В некоторых из них гены в сильной степени репрессированы, в связи с чем проявление тотипотентности становится ограниченным.
Клеточную основу морфогенеза составляет цитодифференцировка. Регенерация растения начинается с вторичной дифференцировки клеток. При этом дедифференцированные клетки вновь приобретают структуру и функции специализированных. Вторичная дифференцировка каллусных клеток не всегда заканчивается морфогенезом и регенерацией растения. Иногда она приводит только к образованию тканей (гистодифференцировка). Таким путем каллусная клетка может превращаться во флоэмные или ксилемные элементы. Другим примером вторичной дифференцировки может служить превращение дедифференцированной активно пролиферирующей клетки в старую неделящуюся каллусную клетку (стационарная фаза роста).
Из всех видов вторичной дифференцировки наибольший интерес представляет морфогенез, так как он позволяет получать целое растение из каллусной клетки. Как отмечалось выше, в основе дифференцировки и морфогенеза лежит последовательное включение различных генов, т.е. дифференцировка клеток определяется дифференциальной активностью генов. Изменение активности структурных генов может быть связано сих дерепрессией, репрессией или амплификацией. Большую роль в этом процессе играют фитогормоны.
Морфогенезом в культуре каллусных тканей можно управлять. На способность изолированных растительных клеток к морфогенезу оказывают влияние как внутренние, так и внешние факторы. К внутренним факторам относятся: видовая принадлежность исходного растения, орган, из которого взят эксплант, возраст экспланта, и даже его массы. В этом случае можно говорить об «эффекте минимальной массы», который сводится к тому, что способность уже детерминированных клеток к дальнейшей дифференцировки зависит от наличия некоторой минимальной массы, необходимой для морфогенеза.
Любопытны работы по выявлению зависимости регенерации растений от скорости их развития. Раннеспелые сорта характеризуются более низким уровнем регенерации по-сравнению с позднеспелыми культурами. Возможно, что выделенные для культивирования in vitro из более быстро развивающихся растений органы и ткани могут иметь жолее короткий период существования инициальных меристематических клеток, обеспечивающих морфологическую компетентность у потенциальных эксплантов.
К внешним факторам, прежде всего, относятся: состав питательной среды, температура, свет (интенсивность и длина фотопериода). Наиболее мощным индуктором морфогенеза, который принято называть стимулом или сигналом морфогенеза, является изменение соотношения между цитокининами и ауксинами, входящими в состав питательных сред.