Клеточная биотехнология
Эффект, вызываемый действием одних и тех же фитогормонов, может быть различным в зависимости от физиологической характеристики ткани-мишени. Её компетентность определяется степенью дифференцировки клеток.
Переход клетки in vitro из дифференцированного состояния к дедифференцировке и активным клеточным делениям обусловлен изменением активности генов (эпигеномной изменчивостью). Активирование одних генов и репрессирование других приводит к изменению в белковом составе клеток. В каллусных клетках появляются специфические белки и одновременно исчезают или уменьшаются в количестве белки, характерные для фотосинтезирующих клеток листа. У двудольных растений процесс репрессии и дерепрессии генов, лежащий в основе дедифференцировки, происходит легче, чем у однодольных. При переходе дедифференцированной клетки к неорганизованному анархическому размножению, приводящему к образованию каллусной ткани, в клетках происходят биохимические и цитологические изменения. Дедифференцировка начинается с использования запасных веществ и разрушения специализированных клеточных органелл. Через 6—12 ч после индукции дедифференцировки клеточная оболочка разрыхляется и разбухает, увеличивается число свободных рибосом, возрастает число элементов аппарата Гольджи, увеличиваются размеры и число ядрышек. Все эти изменения предшествуют началу делений, которые начинаются через 48-72 часа.
Каллусная клетка имеет свой цикл развития и повторяет развитие любой клетки, включая деление, растяжение и дифференцировку, после чего наступает старение и отмирание клетки. Каллусную дифференцировку можно назвать вторичной, но её не следует путать с вторичной дифференцировкой клетки, лежащей в основе морфогенеза. Для того чтобы не произошло старения, утраты способности к делению и отмирание каллусных клеток, первичный каллус, возникший на эксплантах, через 4-6 недель переносят на свежую питательную среду – пассируют. При регулярном пассировании способность к делению может поддерживаться в течение нескольких лет.
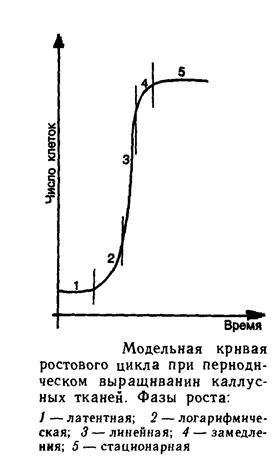
Ростовая кривая каллусных клеток имеет S-образную форму (рис.1). Данный график включает пять фаз. Во время первой – латентной фазы увеличения числа и массы клеток не происходит. Клетки в этот период подготавливаются к делению. Вторая фаза - период экспоненциального роста, характеризующаяся наибольшей митотической активностью и увеличением массы каллусной культуры. Кроме того, рост здесь происходит с ускорением. Третья фаза – линейная, где скорость роста клеток относительно постоянна. Далее наступает фаза замедленного роста, при которой митотическая активность клеток резко снижается. И пятая фаза – стационарная или период деградации. Скорость нарастания клеточной массы здесь равна нулю.
На электронно-микроскопических фотографиях показана тонкая структура молодой, растущей и стареющей клетки каллусной ткани:

а – молодая
б - растущая
в - стареющая
Успех в применении культуры клеток и тканей в первую очередь зависит от оптимизации физиологических процессов, обеспечивающих нормальную пролиферацию, их дифференцировку и регенерацию из них взрослых особей. Наиболее сложной является регенерация растений из отдельных клеток. В первую очередь это касается злаковых растений. Поэтому важнейшее значение имеет выяснение механизма морфогенеза in vitro, регенерация и лежащих в их основы процессов.
Каллусные клетки in vitro сохраняют многие физиолого-биохимические черты, свойственные нормальным клеткам, входящим в состав растительного организма. Каллусные клетки сохраняют способность к синтезу вторичных метаболитов. Морозостойкость и способность к закаливанию присущи каллусным клеткам, полученным от морозостойких растений. Этим свойством не обладают каллусные ткани, полученные от тропических и субтропических культур. Таким образом, устойчивость к низким температурам сохраняется при переходе клетки к каллусному росту. Каллусным тканям свойственна и фотопериодическая реакция, что связано с сохранением активности фитохромов. Общим у каллусные и нормальных клеток растения является и еще ряд признаков, в частности, устойчивость к действию высоких температур, осмотически активных веществ, засолению.
Вместе с тем каллусные клетки обладают отдельными свойствами, отличающими их от нормальных. В них появляются специфические белки, и уменьшается количество белков, характерных для фотосинтезирующих клеток листа, или они совсем исчезают. Каллусные клетки отличаются большой генетической гетерогенностью и физиологической асинхронностью.
В результате выхода из-под контроля организма рост каллусных клеток происходит неорганизованно, асинхронно, и является неограниченным.
Клеточный цикл у каллусных клеток более длительный, чем у растений, произрастающих в открытом грунте. Особенностью каллусных клеток является гетерогенность по возрасту. В каллусной ткани одновременно присутствуют клетки молодые в G1-фазе, старые в G2- и S-фазах цикла клеточных делений.
Значительные отличия наблюдаются в энергетическом обмене каллусных клеток. Они потребляют меньше кислорода по сравнению с нормальными.
Митохондрии в каллусных клетках так же, как и в меристематических, являются слабо развитыми, в них мало крист, что не может не оказывать влияния на активность аэробного дыхания.
Наряду с изменением характера дыхания в каллусных клетках в направлении усиления бескислородного расщепления углеводов происходит также сдвиг в сторону пентозофосфатного пути, который является источником пентоз, необходимых для делящихся клеток.
Длительное время считали, что каллусные клетки генетически строго однородны. Однако клетки каллусной ткани обладают выраженной генетической гетерогенностью. Генетическая неоднородность каллусных клеток выражается, прежде всего, в различной плоидности, т.е. каллусные клетки отличаются по числу хромосом. Генетически стабильными in vitro являются меристематические ткани. В каллусных и суспензионных культурах встречаются клетки, имеющие диплоидный набор хромосом, свойственный исходному растению, полиплоидные клетки, содержащие 3, 4, 5 и более хромосомных наборов. Наряду с полиплоидией в культуре каллусных тканей можно нередко наблюдать анеуплоидию (возрастание или уменьшение хромосомного набора на несколько хромосом). Чем длительнее культивировать каллусные клетки, тем больше они различаются по плоидности. В каллусных клетках табака через четыре года культивирования совсем не остается диплоидных клеток: все клетки становятся полиплоидными или анеуплоидными. Этот факт указывает на то, что изменение плоидности происходит под влиянием условий культивирования и, прежде всего входящих в состав питательной среды веществ. Однако можно интерпретировать его и иначе. Полиплоидные клетки имеют меньшую лаг-фазу и поэтому быстрее переходят к делениям, чем диплоидные. Вследствие этого они и получают преимущество в дальнейших пассажах. Скорее всего, влияние оказывают обе причины.