Клеточная биотехнология
Анатомической дифференцировке предшествует биохимическая дифференцировка, когда видимых различий между клетками мало, но они, не одинаковы по содержанию тех или иных веществ. Удобнее следить за дифференциальной эспрессией генов: появлением новых или снижением уровня старых мРНК и белков. Эти данные позволяют зарегистрировать различия между клетками раньше, чем они станут видимыми на анатомическом уровне. Таким образом, дифференцировка начинается с изменения активности генома, экспрессии одних генов и подавления активности других.
При таком подходе делящиеся клетки меристемы придется считать дифференцированными, так как для прохождения клеточного цикла нужна определённая активность генома, которая и будет отличать эти клетки от других. Анатомы давно обратили внимание на неоднородность клеток меристемы. Можно сказать, что апикальная меристема корня дифференцирована на каллиптроген (инициали чехлика), дерматоген (инициали эпидермальной ткани), инициали коры, покоящийся центр и инициали осевого цилиндра. Для каждой из групп делящихся клеток характерны определенная локализация, направление веретена делений и тип производных клеток. Исследование меристемы методами молекулярной генетики показывает, что обнаруженная анатомами дифференцировка меристемы на зоны совпадает с зонами дифференциальной экспрессией определенных генов. Более того, саму меристему в целом можно достаточно четко выделить по зонам дифференциальной экспрессии. Таким образом, меристема является биохимически дифференцированной тканью.
Дифференциальная экспрессия генов — фундаментальное проявляение дифференцировки, и, как это ни парадоксально, недифференцированных клеток вообще не существует. Понятие «недифференцированный» хорошо работает только там, где в соответствии с задачами исследования исходные различия между клетками не учитывают (или нет методов их обнаружить).
Генетический анализ процесса развития предполагает его разложение на ряд промежуточных этапов, каждый из которых контролируется определённой генетической системой. Развитие есть результат совместной, возможно сменяющей друг друга активности двух генетических систем – первичной и вторичной. Под первичной системой понимается генетический контроль, жёстко регламентирующий переход развивающейся системы из одного состояния в другое, а под вторичной генетической регуляцией – способность системы достигать некоторого конечного состояния автоматически или авторегуляторно.
Геноконтролируемые этапы являются критическими периодами в развитии биологической системы, поскольку именно здесь происходят коренные изменения, связанные с формированием морфофункциональной структуры и определение принципов регулирования. В эти периоды создаются предпосылки негеноконтролируемых переходов системы, в которых она сохраняет свои качественные характеристики и свойства, а также демонстрирует низкую чувствительность к внешним и внутренним изменениям условий развития.
Итак, дифференцировкой можно назвать процесс изменения профиля генной активности, приводящий к дальнейшему изменению функции клеток.
Тотипотентность
В эмбриологии животных процесс дифференцировки изображают как сложный «ландшафт», по которому катится «шар». Шар — это символ клетки, дающей начало новому организму. В развилках шар «совершает выбор» и скатывается по одной из нескольких возможных траекторий. Так и клетки, возникшие при делении зиготы, направляются по одному из возможных путей дифференцировки. При этом клетки теряют «морфогенетический потенциал». Все «траектории» заканчиваются в «море», символизирующем смерть организма.
Если в начале пути у «шара» - клетки много потенциальных возможностей, то по мере приближения к «морю» их становится все меньше.
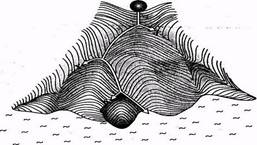
По имени ученого, предложившего такую аналогию, ее называют морфогенетическим ландшафтом Уоддингтона.
Процесс дифференцировки равносилен потере морфогенетического потенциала.
В отличие от клеток животных большинство клеток растений после анатомической дифференцировки легко переходят к делению. Такой процесс называют дедифференцировкой (потерей специализации). При механическом повреждении растения, а также в условиях эксперимента дедифференцировка приводит к образованию каллуса.
Из большинства клеток можно получить новый организм (для клеток животных это невозможно). Практически любая клетка многоклеточного организма содержит полный набор генов, необходимый для формирования организма, однако не каждая клетка может дать начало целому организму. Свойство клетки реализовать имеющуюся генетическую информацию и дать начало целому организму называют тотипотентностью. Тотипотентность клеток растения сравнительно легко реализовать, тогда, как большинство животных клеток не могут образовать новый организм. Таким образом, понятие дифференцировки как снижения морфогенетического потенциала, заимствованное из эмбриологии животных, не применимо к тотипотентным растительным клеткам, так как их морфогенетический потенциал долго остается высоким.
Идея о тотипотентности растительной клетки была выдвинута Г. Хаберландтом еще в 1902 г., хотя и не получила тогда экспериментального подтверждения. Согласно определению Хаберландта, любая клетка растения может дать начало новому организму, и если этого не наблюдается, то только потому, что растительный организм подавляет потенции клетки к развитию. Изоляция клеток от растений способствует проявлению этих потенций.
Культура каллусных тканей и их морфогенетические особенности
Культура изолированных тканей обычно бывает представлена каллусными или опухолевыми тканями. Каллусная культура – это неорганизованная пролиферирующая ткань, состоящая из недифференцированных клеток. В дальнейшем они специализируются как каллусные. Каллус может образовываться как на изолированных участках ткани (эксплантах) in vitro, так и на растении при повреждении.
Каллусная ткань in vitro в основном бывает белого или желтоватого, реже светло-зелёного цвета (полная или зональная пигментация антоцианами). Тёмно-коричневая окраска возникает при старении каллусных клеток и вызвана накоплением в них фенолов. Каллусная ткань аморфна и не имеет конкретной анатомической структуры, но в зависимости от происхождения и условий выращивания она может быть различной консистенции:
1. Рыхлая, состоящая из сильно оводнённых клеток, легко распадающиеся на отдельные агрегаты.
2. Средней плотности, с хорошо выраженными меристематическими очагами.
3. Плотная, в которой дифференцируются элементы камбия и проводящей системы.
Обязательным условием дифференцировки растительной клетки и превращение её в каллусную является присутствие в питательной среде двух групп антагонистических гормонов: ауксинов и цитокининов. Ауксины вызывают процессы дифференцировки клетки, запуская механизмы активизации вторичных мессенджеров, способствующих растяжению клеточных стенок и дальнейшую пролиферацию, а цитокинины вызывают деление уже дифференцированных клеток. Для того чтобы дифференцированные клетки вновь приобрели способность к делению, необходим «возврат» к меристематическому состоянию (дедифференцировка). Размножение дифференцированных клеток приводит к анархическому, неорганизованному росту, в результате чего образуется каллусная ткань. Таким образом, превращение специализированной клетки в каллусную связано с индукцией митозов, способность к которому была потеряна в процессе дифференцировки.