Строение и принцип действия переносчиков
Несколько про - и эукариотических ион-переносящих АТРаз составляют единое семейство и обладают сходными аминокислотными последовательностями и механизмами переноса ионов (табл.2).
Таблица 2.

Наиболее полно охарактеризованы Nа+ /К+-АТРаза из плазматической мембраны животных клеток и Са2+ - АТРаза из саркоплазматического ретикулума. Большинство ферментов этой группы представляют собой единый полипептид с мол. массой 100000; исключение составляет Na+/К+ - АТРаза, выделенная из нескольких источников, которая содержит вторую, меньшую субъединицу с неизвестной функцией. Эти переносчики ингибируются ванадатом и прямо фосфорилируются АТР с образованием фосфорилированного интермедиата, играющего важную роль в транспорте (см. рис.3).
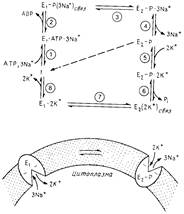
Рис.3 Кинетическая схема для Na+ /К+ - АТРазы. Для Са2+ - АТРазы можно использовать тот же механизм, за исключением того, что вслед за лефосфорилированием Е2 - Р переносчик возвращается в конформацию E1 в незагруженном состоянии (пунктирная линия).
Катионные переносчики этой группы значительно различаются по ионной специфичности (см. табл.2). Неодинакова и стехиометрия транспорта. Например, Са2+ - АТРаза переносит 2Са2+/АТР в полость саркоплазматического ретикулума, в то время как Na+/К+ - АТРаза переносит 3Na+ наружу и 2К + в цитоплазму через плазматическую мембрану. При этом различия в работе АТРазы касаются не только стехиометрии и природы переносимых ионов, но также и того, что Са2+ - АТРаза способна переносить ионы лишь в одном направлении, в то время как Na+ /K+ - АТРаза делает это в обоих направлениях.
Название "фермент Е1E2-типа" было введено в работе, посвященной Na+/К+-АТРазе. Как показали исследования, этот белок существует по меньшей мере в двух различающихся конформациях, для которых характерны разное связывание субстратов и неодинаковая подверженность мягкому протеолизу. Форма Е1 соответствует конформации, в которой места связывания ионов обращены в сторону цитоплазмы (высокое сродство к Na+, низкое - к К+) и которая обладает высоким сродством к АТР. Места связывания ионов в фосфорилированной форме Е2 обращены наружу (высокое сродство к К +, низкое-к Na +). На рис.3 изображен транспортный цикл, в котором участвуют две ненагруженные формы переносчика и две нагруженные, Е2 (2К+) связи E1 (3Na+) связь со "спрятанными" внутри насосного комплекса ионами. Изучение связывания К+ фосфорилированной формой переносчика (Е2-Р) показало, что оно происходит в двух разных местах.
Основные особенности каталитического цикла.
1. E1-форма связывает три иона Na+ с цитоплазматической стороны мембраны и затем взаимодействует с АТР, образуя фосфорилированиый фермент. Фосфорилируется при этом специфический аспартат, консервативный в этой группе ферментов.
2. После отсоединения ADP ионы оказываются "спрятанными" внутри комплекса.
3. Фосфорилирование белка стабилизирует конформацию с низким сродством к Na+; при этом места связывания ионов обращены наружу. Это способствует переходу E1-P в E2-P, в результате которого и осуществляется перенос.
4. В форме Е2-Р места связывания ионов обращены во внеклеточную среду; эта конформация обладает высоким сродством к К+, который связывается, катализирует дефосфорилирование и остается "спрятанным" внутри комплекса. Обратите внимание, что Na+ необходим для быстрого фосфорилирования, а К+ - для быстрого дефосфорилирования. Ванадат связывается с формой Е2, возможно, как некий аналог переходного состояния фосфата. У других ферментов E1E2-типa, например Са2+ - АТРазы, форма Е2-Р дефосфорилируется и переходит в форму E1, которая в свою очередь переходит в незагруженную форму.
5. Лимитирующей стадией каталитического цикла является, по всей вероятности, освобождение К+ и переход его из связанного с ферментом состояния в свободное. Этот процесс стимулируется связыванием АТРс сайтом, обладающим низким сродством.
Следовательно, АТР выполняет две разные функции, выступая в качестве субстрата и аллостерического эффектора. Сколько мест связывания АТР имеет фермент, пока неясно.
Определена аминокислотная последовательность нескольких АТРаз E1E2-типа, включая Na+/К+ - АТРазу (а-субъединица) из нескольких источников, Са2+ - АТРазу, Н+ - АТРазу из плазматической мембраны дрожжей и Neurospora crassa и К+ - АТРазу из S. faecalis. Исходя из профилей гидрофобности, были построены модели, согласно которым эти белковые комплексы содержат 6, 8 или 10 трансмембранных α-спиральных сегментов (рис.4).
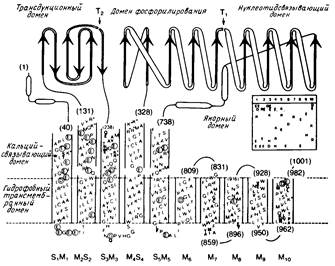
Рис.4
Некоторые участки полипептидов, в том числе и сегмент, содержащий сайт фосфорилирования, в значительной степени гомологичны. Все белки имеют большую гидрофильную петлю, содержащую домены, с которыми, по всей вероятности, связываются нуклеотиды и где происходит фосфорилирование.
У Са+ - АТРазы один расщепляемый трипсином сайт, чувствительный к конформационному переходу Е1 → Е2, находится в "трансдукционном" домене (рис.4). Исследовалось также связывание Са2+ с ферментом; было высказано предположение, что транспорту Са2+ предшествует связывание двух ионов Са2+ с определенными участками внутри белкового комплекса. По всей вероятности, у Са2+ - АТРазы места связывания Са2+ с высоким сродством располагаются на значительном удалении от места связывания нуклеотидов, однако эту гипотезу нужно еще проверить. Основные особенности строения Са2+ - АТРазы, представленные на рис.4, согласуются с данными электронной микроскопии, согласно которым этот белковый комплекс сильно выступает из биослоя в цитоплазму. Вероятно, в условиях in vivo АТРазы Е1 / Е2 - типа агрегируют, образуя по меньшей мере димеры, но подтвердить данное предположение экспериментально очень трудно. Тем не менее очевидно, что мономеры также способны к катализу, по крайней мере в некоторых случаях.
Определена аминокислотная последовательность меньшей α-субъединицы Na+/К+ - АТРазы. Было высказано предположение, что эта субъединица имеет одну или четыре трансмембранные а-спирали.
В заключение отметим, что благодаря легкости клонирования этих мембранных АТРаз и возможности использования разных экспериментальных подходов эти системы являются отличным объектом для применения к ним направленного мутагенеза и других генетических методов. Подобные методы уже применяются в исследованиях F1F0 - АТРаз.
6.2 АТР-азы F1F0-типа из митохондрий, хлоропластов и бактерии
Большинство бактерий, а также митохондрии и хлоропласты содержат родственные АТРазы F1F0 - типа, которые используют трансмембранный протонный электрохимический градиент для синтеза АТР из ADP и неорганического фосфата. В физиологических условиях эти ферменты являются АТР-синтазами. Они содержат от 8 (е. coli) до 13 (митохондрии сердца быка) различных субъединиц и, таким образом, являются гораздо более сложными структурами, чем АТРазы E1E2-типa из плазматических мембран. АТРазы F1F0-типа состоят из двух частей: