Механизмы реализации путей гибели клетки
Постэмбриональный гемопоэз. Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови (клеточное обновление), который компенсирует физиологическое разрушение дифференцированных клеток.
Миелопоэз происходит в миелоидной ткани (textus myeloideus), расположенной в эпифизах трубчатых и полостях многих губчатых костей.
Здесь развиваются форменные элементы крови: эритроциты, гранулоциты, моноциты, кровяные пластинки, предшественники лимфоцитов.
В миелоидной ткани находятся стволовые клетки крови и соединительной ткани.
Предшественники лимфоцитов постепенно мигрируют и заселяют такие органы, как тимус, селезенка, лимфатические узлы и др.
Лимфопоэз происходит в лимфоидной ткани (textus lymphoideus), которая имеет несколько разновидностей, представленных в тимусе, селезенке, лимфатических узлах. Она выполняет основные функции: образование Т - и В-лимфоцитов и иммуноцитов (плазмоцитов и др.).
Миелоидная и лимфоидная ткани являются разновидностями соединительной ткани, т.е. относятся к тканям внутренней среды. В них представлены две основные клеточные линии - клетки ретикулярной ткани и гемопоэтические.
Ретикулярные, а также жировые, тучные и остеогенные клетки вместе с межклеточным веществом (матрикс) формируют микроокружение для
гемопоэтических элементов. Структуры микроокружения и гемопоэтические
клетки функционируют в неразрывной связи. Микроокружение оказывает
воздействие на дифференцировку клеток крови (при контакте с их рецепторами или путем выделения специфических факторов).
Для миелоидной и всех разновидностей лимфоидной ткани характерно
наличие стромальных ретикулярных и гемопоэтических элементов,
образующих единое функциональное целое. В тимусе имеется сложная строма, представленная как соединительнотканными, так и ретикулоэпителиальными клетками. Эпителиальные клетки секретируют особые вещества - тимозины, оказывающие влияние на дифференцировку из СКК Т-лимфоцитов. В лимфатических узлах и селезенке специализированные ретикулярные клетки создают микроокружение, необходимое для пролиферации и дифференцировки в специальных Т - и В-зонах Т - и В-лимфоцитов и плазмоцитов.
СКК являются плюрипотентными (полипотентными) предшественниками всех клеток крови и относятся к самоподдерживающейся популяции клеток. Они редко делятся. Впервые представление о родоначальных клетках крови сформулировал в начале XX в.А. А. Максимов, который считал, что по своей морфологии они сходны с лимфоцитами. В настоящее время это представление нашло подтверждение и дальнейшее развитие в новейших экспериментальных исследованиях, проводимых главным образом на мышах. Выявление СКК стало возможным при применении метода колониеобразования.
Экспериментально (на мышах) показано, что при введении смертельно облученным животным (утратившим собственные кроветворные клетки) взвеси клеток красного костного мозга или фракции, обогащенной СКК, в селезенке появляются колонии клеток - потомков одной СКК. Пролиферативную активность СКК модулируют колониестимулирующие факторы (КСФ), интерлейкины (ИЛ-3 и др.). Каждая СКК в селезенке образует одну колонию и называется селезеночной колониеобразующей единицей (КОЕ-С).
Подсчет колоний позволяет судить о количестве стволовых клеток, находящихся во введенной взвеси клеток. Таким образом, было установлено, что у мышей на 105 клеток костного мозга приходится около 50 стволовых клеток, из селезенки - 3,5 клетки, среди лейкоцитов крови - 1,4 клетки.
Исследование очищенной фракции стволовых клеток с помощью электронного микроскопа позволяет считать, что по ультраструктуре они очень близки к малым темным лимфоцитам.
Исследование клеточного состава колоний позволило выявить две линии их дифференцировки. Одна линия дает начало мультипотентной клетке - родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (КОЕ-ГЭММ). Вторая линия дает начало мультипотентной клетке - родоначальнице лимфопоэза (КОЕ-Л). Из мультипотентных клеток дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные родоначальные (прогениторные) клетки.
Методом колониеобразования определены родоначальные унипотентные клетки для моноцитов (КОЕ-М), нейтрофильных гранулоцитов (КОЕ-Гн), эозинофилов (КОЕ-Эо), базофилов (КОЕ-Б), эритроцитов (БОЕ-Э и КОЕ-Э), мегакариоцитов (КОЕ-МГЦ), из которых образуются клетки-предшественники (прекурсорные). В лимфопоэтическом ряду выделяют унипотентные клетки - предшественницы для В-лимфоцитов и соответственно для Т - лимфоцитов. Полипотентные (плюрипотентные и мультипотентные), олигопотентные и унипотентные клетки морфологически не различаются.
Все приведенные выше стадии развития клеток составляют четыре основных компартмена: I - стволовые клетки крови (плюрипотентные, полипоте нтные); II - коммитированные родоначальные клетки (мультипотентные); III - коммитированные родоначальные (прогенторные) олигопотентные и унипотентные клетки; IV - клетки-предшественники (прекурсорные).
Дифференцировка полипотентных клеток в унипотентные определяется действием ряда специфических факторов - эритропоэтинов (для эритробластов), гранулопоэтинов (для миелобластов), лимфопоэтинов (для лимфобластов), тромбопоэтинов (для мегакариобластов) и др.
Из каждой клетки-предшественницы происходит образование конкретного вида клеток. Созревание каждого вида клеток проходит ряд стадий, которые в совокупности образуют компартмент созревающих клеток (V).
Зрелые клетки представляют последний компартмент (VI). Все клетки V и VI компартментов морфологически можно идентифицировать.

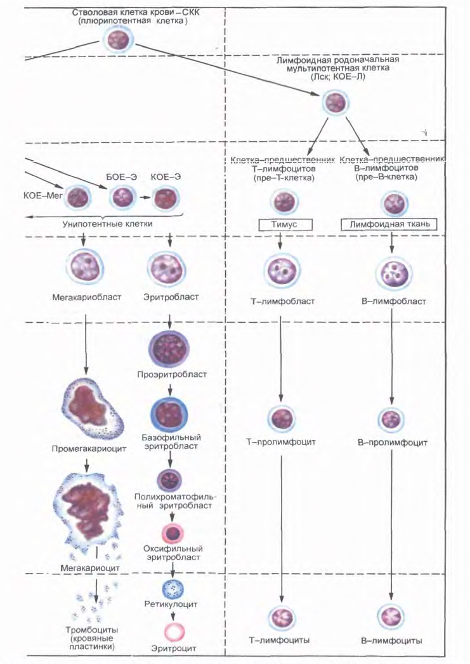
Рис.18. Постэмбриональный гемопоэз, окраска азур 11-эозином (схема по НАЮриной). Стадии дифферениировки крови: I-IV - морфологически неидентифицируемые клетки; V - VI - морфологически идентифицируемые клетки. Б - базофил; БОЕ - бурстобраэуюшая единица; Г - гранулоциты; Гн - гранулоцит нейтрофильный; КОЕ - колониеобразующ! единицы; КОЕ-С - селезеночная колониеобразующая единица; Л - лимфоцит; Лек - mt фоидная стволовая клетка; М - моноцит; Мет - мегакариошгг; Эо - эозинофил; Э - эритроцит.
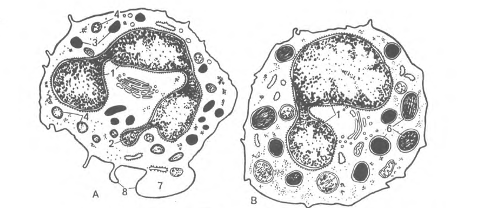
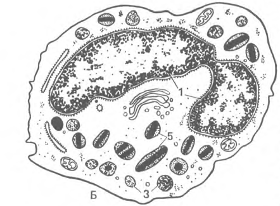
Рис. 19. Ультрамикроскопическое строение гранулоцитов (схема по НА. Юриной и Л.С. Румянцевой).
А - сегментоядерный нейтрофильный гранулоцит; Б – эозинофильный (ацидофильный) гранулоиит; В - базофильный фанулоцит: 1 - сегменты ядра; 2 - тельце полового хроматина; 3 – первичные (азурофильные) гранулоциты; 4 - вторичные (специфические) гранулы; 5 - зрелые специфические гранулы эозинофила, содержащие кристаллоиды; б - гранулы базофила различной величины и плотности; 7 - периферическая зона, не содержащая органелл; 8 – микроворсинки и псевдоподии.