Механизмы реализации путей гибели клетки
По содержанию агглютиногенов и агглютининов различают 4 группы крови: в крови А (0) группы отсутствуют агглютиногсны А и В, но имеются а - и р-агтлютинины; в крови А (П) группы имеются агглютиноген А и р-агглютинин; в крови В (Ш) группы содержатся В-агглютиноген и а-агглютинин; в крови AB (IV) группы имеются агглютиногены А и В и нет агглютининов. При переливании крови для предотвращения гемолиза (разрушение эритроцитов) нельзя допускать вливания реципиентам эритроцитов с агглютиногенами А или В, имеющим а - или р-агглютинины. Поэтому лица с (0) A группой крови являются универсальными донорами, т.е. их кровь может быть перелита всем людям с другими группами крови.
Соответственно лица с AB (FV) группой крови являются универсальными реципиентами, т.е. им можно перелить любую группу крови.
На поверхности эритроцитов имеется также резус-фактор (Rh-фактор) - агглютиноген. Он присутствует у 86 % людей; у 14 % отсутствует (резус-отрицательные). Переливание резус-положительной крови резус-отрицательному пациенту вызывает образование резус-антител и гемолиз эритроцитов. Агглютинация эритроцитов свойственна нормальной свежей крови, при этом образуются так называемые "монетные столбики".
Это явление связано с потерей заряда плаэмолеммы эритроцитов. Скорость оседания (агглютинации) эритроцитов (СОЭ) в 1 ч у здорового человека составляет 4-8 мм у мужчин и 7-10 мм у женщин. СОЭ может значительно изменяться при заболеваниях, например при воспалительных процессах, и поэтому служит важным диагностическим признаком. В движущейся крови эритроциты отталкиваются из-за наличия на их плазмолемме одноименных отрицательных зарядов. Поверхность плазмолеммы одного эритроцита составляет около 130 мкм2 крови. При окрашивании мазка крови азур И-эозином по Романовскому - Гимзе большинство эритроцитов приобретают оранжево-розовый цвет (оксифильны), что обусловлено высоким содержанием в них гемоглобина.
В небольшой части эритроцитов A-5 %), являющихся более молодыми формами, сохраняются остатки органелл (рибосомы, гранулярный, эндоплазматический ретикулум), которые проявляют базофилию. Такие эритроциты окрашиваются как кислыми красителями (эозин), так и основными (азур II) и называются полихроматофилъными. При специальной суправитальной окраске (бриллиант-крезилфиолетовым) в них выявляются зернисто-нитчатые структуры, поэтому их называют ретикулоцитами. Эритроциты различаются по степени насыщенности гемоглобином.
Среди них выделяются нормохромные, гипохромные и гиперхромные, соотношение между которыми значительно изменяется при заболеваниях. Количество гемоглобина в одном эритроците называют цветным показателем. Электронно-микроскопически гемоглобин выявляется в гиалоплазме эритроцита в виде многочисленных плотных гранул диаметром 4-5 нм.
Гемоглобин - это сложный белок F8 КД, состоящий из 4 полипептидных цепей глобина и гема (железосодержащий порфирин), обладающий высокой способностью связывать кислород. В норме у человека содержится два типа гемоглобина - НЬА и HbF. Эти гемоглобины различаются составом аминокислот в глобиновой (белковой) части.
У взрослых людей в эритроцитах преобладает НЬА, (от англ. adult - взрослый), составляя 98 %. Он содержит две а-глобиновые цепи и две C-глобиновые цепи, включающие 574 аминокислоты.
HbF, или фетальный гемоглобин (от англ. foetus - плод), составляет у взрослых около 2 % и преобладает у плодов. К моменту рождения ребенка HbF составляет около 80 %, а НЬА только 20 %. Эти гемоглобины отличаются составом аминокислот в глобиновой (белковой) части.
Железо (Fe2+) в геме может присоединять О2 в легких (в таких случаях образуется оксигемоглобин - НЬО2) и отдавать его в тканях путем диссоциации НЬО, на кислород (О2) и НЬ; валентность Fe2+ не изменяется.
При ряде заболеваний (гемоглобинозы, гемоглобинопатии) в эритроцитах появляются другие виды гемоглобинов, которые характеризуются изменением аминокислотного состава в белковой части гемоглобина.
В настоящее время выявлено более 150 видов аномальных гемоглобинов. Например, при серповидно-клеточной анемии имеет место генетически обусловленное повреждение в C-цепи гемоглобина - глютаминовая кислота, занимающая 6-е положение в полипептидной цепи, заменена на аминокислоту валин. Такой гемоглобин обозначается как HbS (от англ. sickle - серп), так как в условиях понижения парциального давления О2 он превращается в тектоидное тело, придавая эритроциту форму серпа. В ряде стран тропического пояса определенный контингент людей являются гетерозиготными для серповидных генов, а дети двух гетерозиготных родителей по законам наследственности дают либо нормальный тип B (5%), либо бывают гетерозиготными носителями, и 25 % страдают серповидноклеточной анемией.
Гемоглобин способен связывать О2 в легких, при этом образуется оксигемоглобин, который транспортируется ко всем органам и тканям и там отдает О2. В тканях выделяемая СО поступает в эритроциты и соединяется с НЬ, образуя карбоксигемоглобин. При разрушении эритроцитов (старых или при воздействии различных факторов - токсины, радиация и др.) гемоглобин выходит из клеток, и это явление называется гемолизом. Старые эритроциты разрушаются макрофагами главным образом в селезенке, а также в печени и костном мозге, при этом НЬ распадается, а высвобождающееся из железосодержащего гема железо используется для образования новых эритроцитов.
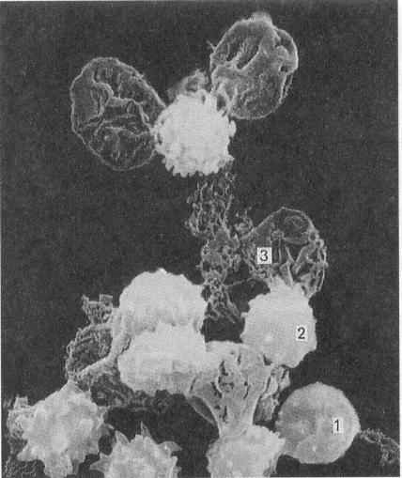
Рис 15. Электронная микрофотография гемолиза эритроцитов и образование их “теней”
В макрофагах НЬ распадается на пигмент билирубин и гемосидерин - аморфные агрегаты, содержащие железо. Железо гемосидерина связывается с трансферрином - негеминовым белком плазмы, содержащим железо, и захватывается специальными макрофагами костного мозга. В процессе образования эритроцитов (эритропоэз) эти макрофаги передают трансферрин в формирующиеся эритроциты, что послужило основанием назвать их клетками-кормилками.
В цитоплазме эритроцитов содержатся ферменты анаэробного гликолиза, с помощью которых синтезируются АТФ и НАДН, обеспечивающие энергией главные процессы, связанные с переносом О2 и СО2, а также поддержание осмотического давления и перенос ионов через плазмолемму эритроцита. Энергия гликолиза обеспечивает активный транспорт катионов через плазмолемму, поддержание оптимального соотношения концентрации К+ и Na+ в эритроцитах и плазме крови, сохранении формы и целостности мембраны эритроцита. НАДН участвует в метаболизме НЬ, предотвращая окисление его в метгемоглобин.
Эритроциты участвуют в транспорте аминокислот и полипептидов, регулируют их концентрацию в плазме крови, т.е. выполняют роль буферной системы. Постоянство концентрации аминокислот и полипептидов в плазме крови поддерживается с помощью эритроцитов, которые адсорбируют их избыток из плазмы, а затем отдают различным тканям и органам. Таким образом, эритроциты являются подвижным депо аминокислот и полипептидов.
Сорбционная способность эритроцитов связана с состоянием газового режима (парциальное давление О2 и СО2 - Ро, Рсо): в частности, при действии О2 наблюдаются выход аминокислот из эритроцитов и увеличение их содержания в плазме.