Страница
9
5.3 Дыхательная система митохондрий
Внутренняя мембрана митохондрий является местом, где осуществляется окислительное фосфорилирование. Суть процесса состоит в сопряжении потока электронов, направленного от органических субстратов к кислороду, с перемещением протонов из матрикса митохондрий через мембрану в межмембранное пространство. Как и предыдущие две электронтранспортные цепи, компоненты этой системы тоже представлены не в эквимолярных количествах, т.е. дыхательная цепь не может функционировать как долгоживущий мультиферментный комплекс. Однако в данном случае число оборотов терминального фермента, цитохром с-оксидазы, весьма велико, а оценить, какая стадия является лимитирующей, в такой ситуации очень непросто. Цепь состоит из четырех трансмембранных мультисубъединичных комплексов, растворимого цитохрома с и убихинона-10. Перенос электронов через комплекс II не сопровождается переносом протонов. Однако реакции, катализируемые комплексом I, комплексом III и комплексом IV, сопровождаются векторным переносом протонов через мембрану. Эти реакции являются электрогенными и приводят к генерации трансмембранного электрического потенциала. Такие ферменты в составе цепи называются "местами сопряжения". Относительно механизмов перемещения протонов пока нет единого мнения, хотя в литературе обсуждаются конкретные модели, в частности для комплексов III и IV. Образующаяся трансмембранная разность электрохимических потенциалов протонов уменьшается за счет работы протонного канала АТР-синтазы. Энергия потока используется этим ферментом для синтеза АТР.
Концентрация компонентов дыхательных цепей в митохондриальной мембране довольно высока, однако существование эквимолярных комплексов между образующими эти цепи мультисубъединичными ферментами нельзя считать доказанным.
В литературе обсуждаются модели электронного транспорта с участием таких переходных долгоживущих белковых агрегатов, однако имеющиеся кинетические данные можно объяснить, не предполагая образование таких суперкомплексов. Цитохром с выполняет функцию челнока, быстро переносящего электроны между комплексами III и IV, аналогично адренодоксину в содержащей цитохром Р450 системе митохондрий. При физиологической ионной силе цитохром с может диффундировать не только вдоль поверхности бислоя, но и в объеме раствора, что увеличивает его способность к быстрому переносу электронов. Данная система обладает рядом особенностей, не характерных для описанных выше примеров электронтранспортных цепей.
Основные компоненты дыхательной цепи организованы в не-диссоциирующие комплексы. Например, комплекс III состоит из нескольких субъединиц и содержит три гема и один железосерный центр. Перенос электронов между простетическими группами внутри каждого из компонентов происходит быстро и не требует их случайных столкновений.
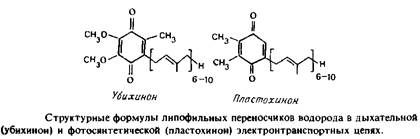
Жирорастворимые переносчики водорода служат для переноса восстановительных эквивалентов не только между ферментами, но и с одной стороны бислоя на другую. Показано, что убихинон обладает способностью к быстрой латеральной диффузии в плоскости мембраны, хотя вопрос о величине коэффициента диффузии не решен. В модельных системах убихинон также может переносить восстановительные эквиваленты через бислой. Убихинон при перемещениях не выходит из мембраны, однако в ходе редокспревращений он может захватывать или высвобождать протоны на границе раздела фаз липид-вода с любой стороны мембраны. Кинетические свойства изолированных ферментов, использующих убихинон в качестве субстрата, можно с успехом анализировать, используя уравнения Михаэлиса-Ментен с соответствующими значениями Км и Итах, если известна реальная концентрация хинона в бислое. Иное дело, если мы рассматриваем стационарную кинетику систем, в которых одна популяция ферментов сопряжена с другой посредством свободно диффундирующего пула хинонов как интермедиатов. Для описания подобных систем приходится применять специальные уравнения. Тот факт, что компоненты дыхательной цепи связаны через свободно диффундирующий пул хинонов, не вызывает сомнений, но в вопросе, является ли диффузия хинона лимитирующей стадией в работе электронтранспортной цепи, единого мнения среди исследователей нет.
Протоны, перенесенные через мембрану при работе дыхатель- ной цепи, возвращаются обратно с помощью АТР-синтазы, замыкая тем самым протонный цикл. Механизм, по которому протоны переносятся от компонентов электронтранспонтной цепи к АТР-синтазе, тоже до конца не установлен, и этот вопрос вряд ли будет разрешен в ближайшем будущем. Большинство исследователей считают, что протоны, переносимые через бислой, быстро приходят в равновесие со всей водной фазой. Согласно другой точке зрения, существует локализованный протонный поток и протоны переносятся либо вдоль поверхности бислоя, либо внутри бислоя, либо прямо на АТР-синтазу. Большинство данных в пользу этого механизма носят косвенный характер, но в литературе имеются свидетельства быстрого перемещения протонов вдоль поверхности фосфолипидного монослоя в нефизиологических условиях. Возможно, при некоторых обстоятельствах такой локализованный протонный поток имеет какое-то значение.
Таким образом, данная электронтранспортная система представляет собой совокупность небольшого числа высокоорганизованных комплексов, связанных между собой низкомолекулярными подвижными переносчиками, как липофильными, так и водорастворимыми. Кинетику переноса электронов можно объяснить в рамках модели свободно диффундирующих форм, которые могут перемещаться вдоль мембраны на расстояние более 100 А.
5.4. Фотосинтетическая электронтранспортная система тилакоидов
К фотосинтетической электронтранспортной системе можно также отнести светособирающие комплексы, поглощающие свет и передающие энергию электронного возбуждения на два реакционных центра. Захват энергии возбуждения реакционными центрами приводит к разделению зарядов и образованию на противоположных сторонах мембраны сильного окислителя и сильного восстановителя. Это в свою очередь Ведет к окислению воды до молекулярного кислорода фотосистемой II и создает движущую силу для транспорта электронов по цепи, сопряженного с трансмембранным переносом протонов. Образующийся на бислое градиент электрохимических потенциалов протонов используется для синтеза АТР с помощью АТР-синтазы, называемой сопрягающим фактором. Структура фотосистемы II, по-видимому, очень похожа на структуру бактериального фотореакционного центра, которая была детально изучена методом рентгеноструктурного анализа. &б/-Комплекс аналогичен комплексу III дыхательной цепи митохондрий, а сопрягающий фактор - митохондриальной АТР-синтазе. Роль пластоцианина очень напоминает роль цитохрома с в дыхательной цепи, являющегося растворимым переносчиком электронов, а пластохинон представляет собой полный аналог убихинона в электронтранспортной системе митохондрий.
В отличие от дыхательной цепи основные мембранные комплексы представлены в фотосинтетической системе примерно в эквимо-лярном соотношении. Однако какие-либо серьезные указания в литературе на формирование суперкомплексов отсутствуют. Напротив, для данной системы характерна поразительная латеральная гетерогенность, в результате которой фотосистема II оказывается локализованной в гранальных, плотно упакованных участках, а фотосистема I и сопрягающий фактор - в стромальных участках тилакоида. Пластохинон и комплекс, по-видимому, распределены между этими участками равномерно. Вследствие такого латерального разделения для сопряжения двух фотосистем необходима диффузия на расстояние по крайней мере 1000 А. Скорее всего основным переносчиком восстановительных эквивалентов на такие расстояния является пластохинон, хотя скорость его латериальной диффузии точно не известна. В оптимальных условиях лимитирующей стадией, вероятно, является окисление пластохинона £б/-комплексом, однако связано ли это со скоростью его латеральной диффузии или с работой самого фермента - неясно.