Изучение трехмерной структуры с помощью рентгеновской дифракции и реконструкции изображения
Проекции электронной плотности в двух направлениях недостаточны для построения трехмерной структуры. Поэтому, наклоняя образец, получают проекции образца под разными углами и используют их для реконструкции трехмерного изображения объекта. Таким образом строят карту электронной плотности в мембране на Разных уровнях. Обычно приводят данные о профиле электронной плотности через каждые 15-25 А.
Таблица 2. Мембранные белки, структуру которых определяли методом реконструкции изображения
|
Белок |
Трехмерное разрешение, А |
Ссылки |
|
1. NAPH: убихинон оксидоредуктаза (митохондрии) |
13 |
[ИЗ] |
|
2. Цитохром с-оксидаза (митохондрии) |
20 |
[314, 466] |
|
3. Убихинол-цитохром с - оксидоредуктаза (митохондрии) |
25 |
[843] |
|
4. Свет ос обирающий комплекс, содержащий |
16 |
[795] |
В табл.2 представлен список белков, которые были изучены этим методом. Во всех случаях, кроме бактериородопсина, уровень разрешения был достаточен только для очерчивания общих контуров молекул и определения их размеров.
Однако даже такая информация может быть очень ценной. Например, выяснилось, что многие из этих молекул в двумерных кристаллах существуют в виде отдельных мультимеров, а порин и бактериородопсин являются тримерами.
В случае порина существует четко наблюдаемый канал, в образовании которого на внешней поверхности клетки участвует каждый из трех отдельных полипептидов; сливаясь, эти полипептиды образуют одиночный канал на периплазматической поверхности наружной мембраны Е. coli.
Коннексин выглядит как гексамер, а ацетилхолиновый рецептор - как симметричный пентамер. Ферменты дыхательной цепи митохондрий, убихинолцитохром с-оксидоредуктаза и цитохромоксидаза, являются димерами, хотя неясно, существуют ли эти димеры in vivo. Удивительно, как сильно белки иногда выступают над поверхностью бислоя.
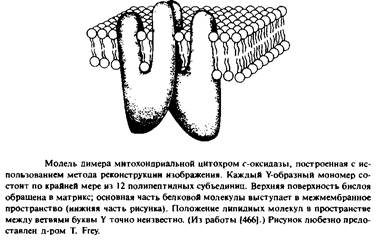
Например, отдельные части цитохром с-оксидазы возвышаются над поверхностью бислоя на 50 А 1314, 466] ; такая же картина наблюдается для ацетилхолинового рецептора. Напротив, порин и бактериородопсин почти не выдаются над поверхностью мембран. На рис.3.3 представлена структура цитохром с-оксидазы, полученная с низким разрешением.
2.2 Три примера структурных исследований мембранных белков
Рассмотрим три примера изучения интегральных мембранных белков, иллюстрирующие большое разнообразие используемых для этого методов. Наиболее известными структурами являются реакционные центры R. viridis и R. sphaeroides, исследование которых с помощью рентгеновской дифракции было весьма успешным. Еще одна наиболее полно изученная структура - бактериородопсин Н. halobium; для его исследования применялся метод реконструкции изображения, а также другие подходы. Порин и родственные белки наружной мембраны Е. соЧ изучали в основном с помощью генетических и молекулярно-биологических методов, позволяющих идентифицировать функционально важные участки.
3. Структура фотосинтетических реакционных центров ft. viridis и ft. sphaeroides
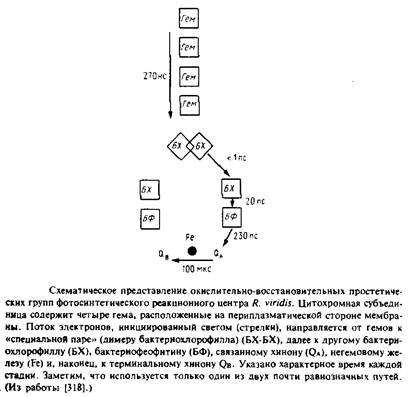
Фотосинтетические реакционные центры представляют собой комплексы белков с пигментами; в них происходит первичное разделение зарядов в фотосинтетических мембранах. Лучше всего охарактеризованы комплексы из пурпурных несерных бактерий; они обычно состоят из трех белковых субъединиц - Н, М и L. Реакционный центр Rhodopseudomonas viridis имеет также четвертую субъединицу - цитохром с-типа. Простетическими группами этого комплекса являются четыре гемогруппы, четыре бактериохлорофилла Ь, два бактериофеофитина, одно негемовое железо, один менахинон и один убихинон. Под действием света электрон переходит от первичного донора электронов, так называемой "специальной пары" - молекул бактериохлорофилла, образующих димер, к бактериофеофитину, а затем к первичному хиноновому акцептору QA. В конце концов электрон восстанавливает вторичный акцептор Qb в ходе реакции, при которой протоны поступают из раствора на восстановленный хинон. Qb находится в равновесии с хинонным пулом в би-слое. Окисленный первичный донор электронов, "специальная пара", восстанавливается цитохромом с-типа. Поскольку цитохром и хинон расположены на противоположных сторонах фотосинтетической мембраны, светозависимый электронный транспорт электроге-нен и генерирует трансмембранную разность потенциалов.
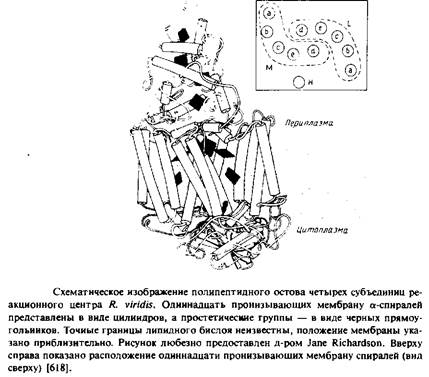
Суммарная мол. масса реакционного центра из R. viridis составляет примерно 150 ООО, а кажущаяся мол. масса субъединиц - 38 ООО, 35 ООО, 28 ООО и 24 ООО. Заметим, что электрофорез в ПААГ-ДСН дает неправильные молекулярные массы для Н, М и L субъединиц; об этом свидетельствуют данные о числе аминокислотных остатков в каждом полипептиде, полученные при секвенировании ДНК. Очищенный комплекс был закристаллизован с использованием сульфата аммония как осаждающего агента в присутствии детергента N. N-диме-тилдодециламин-М-оксида и органического амфифильного соединения гептан-1,2,3-триола. Кристаллы были в достаточной степени упорядочены и фотохимически активны. Структура этого четырехсубъединичного белка была установлена с разрешением около 3 А. Детергент в кристаллах не упорядочен, поэтому невозможно точно определить положения границ погруженных в мембрану участков. Размеры комплекса - 30 х 70 х 130 A. L - и М-субъединицы содержат по пять трансмембранных а-спиральных участков, а у Н-субъединицы такой участок только один.
Итак, все трансмембранные области этого белкового комплекса имеют а-спиральную конфигурацию. Длина каждого а-спирального сегмента составляет примерно 40 А, этого достаточно для пересечения мембраны.
Структура белкового комплекса из R. viridis напоминает сэндвич. L - и М-субъединицы уложены одинаковым образом и расположены в центре сэндвича. Они пересекают бислой и связаны со всеми простетическими группами, за исключением гемов. Сегменты L и М, которые соединяют трансмембранные сегменты по обе стороны мембраны, участвуют в связывании цитохрома и Н-субъединиц. Цитохром образует "шапочку" на наружной поверхности бислоя, а гидрофильная часть субъединицы Н - аналогичную структуру на цитоплазматической поверхности. Трансмембранная а-спираль на N-конце Н-субъединицы контактирует с цитохромом на противоположной стороне мембраны.
Эти межмолекулярные взаимодействия в кристаллах осуществляются между участками Н-субъединицы и цитохромом, которые в норме контактируют с водой. Возможно, эти два белка, образующие гидрофильные "шапочки", способствуют формированию высокоупорядоченных кристаллов.