Страница
2
И лабораторные крысы, и черно-серебристые лисицы, хотя и ведут свое происхождение от соответствующих диких форм, в течение многих поколений разведения в неволе не испытывали действия естественного отбора. Иными словами, в популяциях таких животных не было «выживания наиболее приспособленных», и соответственно доля животных, способных к быстрым адекватным реакциям на меняющиеся внешние условия, оказалась уменьшенной. Отражением этого можно считать снижение доли правильных решений теста на элементарную рассудочную деятельность.
Л. В. Крушинский (1986) предполагал, что в случае прекращения действия естественного отбора при размножении животных в неволе разрушаются сложные полигенные системы (или «коадаптированные комплексы»), которые в естественных условиях обеспечивают приспособление животных (через механизмы поведения) к изменяющимся и часто неблагоприятным внешним условиям.
Среди лабораторных мышей также были обнаружены генетические группы, у которых доля правильных решений задачи достоверно превышала случайную. Это были мыши с робертсоновской транслокацией (слиянием) хромосом Rb(8,}7) llem. В начале этого исследования способность к экстраполяции была проанализирована у значительного числа мышей с различными нарушениями кариотипа, в частности с робертсоновскими транслокациями разных хромосом. У животных с большинством таких мутаций доля правильных решений теста также не отличалась от 50%-го случайного уровня. В то же время мыши, у которых было слияние хромосом 8 и 17 (транслокации Rb(8,17)llem и Rb(8,17)6Sic; оно возникло совершенно независимо и найденно в разных лабораториях), оказались способными к экстраполяции. Рис. 1В показывает, что мыши со слиянием хромосом 8 и 17 (3—5) достоверно решали задачу на экстраполяцию, тогда как мыши с нормальным кариотипом (1,2) и со слиянием других хромосом (6) задачи не решали.
Мыши с этой хромосомной мутацией, в течение более 20 лет разводившиеся в нашей лаборатории, устойчиво показывали отличный от случайного уровень решения задачи на экстраполяцию. Позднее мы исследовали этот вопрос с использованием уникальной генетической модели — мышей 4 инбредных линий, которые попарно различались либо по генотипу (СВА и C57BL/6J), либо по наличию или отсутствию этой транслокации (Полетаева, 1998).
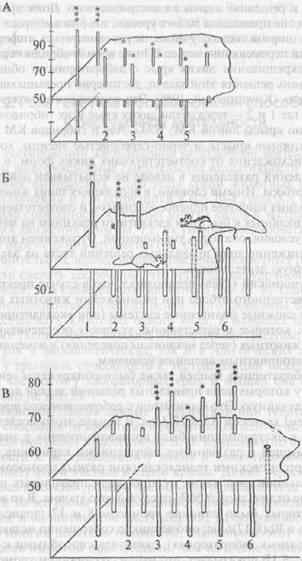
Рис. 1. Успешность решения теста на экстраполяцию животными разных генетических групп (пояснения в тексте).
А — решение теста лисицами; Б — крысами; В — мышами. Высота столбца соответствует доле правильных решений задачи при первом (ближний ряд) и многократных (дальний ряд) предъявлениях задачи.
Эксперименты с животными этих линий показали, что усиление способности к решению теста на экстраполяцию и другие особенности поведения, а также особенности обмена катехола-минов у этих мышей связаны именно с наличием в их кариотипе данной робертсоновской транслокации.
Возможно, что причиной, лежащей в основе этих изменений в функции ЦНС при данной хромосомной перестройке, могут быть изменения в пространственном расположении генетического материала в интерфазном ядре, возникшие как следствие слияния хромосом.
Данные по различиям способности к экстраполяции у животных, отличающихся друг от друга генетически, естественно, не стоят особняком, а являются частью огромной «базы данных», созданной к сегодняшнему дню учеными, работающими в области генетики поведения.
Генетические исследования затрагивают практически все формы поведения, в том числе и способность к обучению, и способность к формированию пространственных предстаапений. Для того чтобы вкратце познакомиться с этим материалом, необходимо сначала дать краткий очерк основных методологических особенностей данного направления. Далее приводятся примеры использования генетических методов для изучения когнитивных способностей животных, а также краткое описание исследований генетических закономерностей психических способностей человека.
3. Методы и объекты генетики поведения.
Генетические подходы к исследованию поведения позволяют выяснить, с чем именно связана изменчивость интересующего нас признака, т.е. в какой степени она связана с изменчивостью генотипов данной группы животных, а в какой — с внешними по отношению к генотипу событиями, воздействующими на ЦНС, а следовательно, и на поведение. В таких исследованиях важную роль играет использование так называемых генетических моделей — групп животных, состоящих из генетически «охарактеризованных», нередко идентичных (или почти идентичных) по генотипу особей с определенными физиологическими или биохимическими особенностями. Используют, например, линейных животных — инбредные и селектированные линии. Между инбредными линиями (мышей или крыс) обнаруживаются различия по тем или иным признакам поведения. Выявление таких межлинейных различий — обычно первый этап исследования. Следующим шагом в классических исследованиях по генетике поведения бывает скрещивание животных из линий, обнаруживших контрастные значения признака, с получением гибридов и анализом расщепления признаков во втором и последующем поколениях. Наряду с этим в генетике поведения используются селектированные линии, сформированные путем искусственного отбора на высокие и низкие значения какого-либо признака поведения (в таких случаях для скрещивания в последовательных поколениях отбираются животные соответственно с высокими и низкими значениями интересующего исследователя признака). После выведения таких линий нередко проводится их скрещивание и анализ проявления признаков у потомства.
Данные такого классического генетического анализа делают возможным вывод и о количестве генов, которые определяют основной вклад в изменчивость изучаемого признака поведения. Тестирование поведения гибридов первого поколения дает информацию о доминантном, промежуточном или рецессивном наследовании интересующего признака. Если данный признак определяется одним, двумя или тремя генами, то это можно установить по картине его распределения у гибридов второго поколения и потомков возвратного скрещивания. Если же в определении признака участвует большее число генов, то необходимо применять методы генетики количественных признаков. Современный этап развития науки обогатил генетику поведения новыми методами. К их числу относятся:
метод рекомбинантных инбредных линий (см.: Nesbitt, 1992);
метод QTL—quantitative trait loci (Le Roy, 1999);
создание и исследование мозаичных и химерных животных (Мак Ларен, 1979);
создание трансгенных организмов и животных-нокаутов (см.: Jones, Mormede, 1999).
Нейрогенетика и генетика поведения сформировались в большой степени благодаря работам на дрозофиле (Drosophila melanogester). Это относится и к генетическому исследованию процесса развития нервной системы, и к выявлению специфических для нервной системы генов и генных комплексов, оказывающихся сходными и у дрозофилы, и у млекопитающих.
Мыши (Mus musculus) также чрезвычайно важный экспериментальный объект нейрогенетики и генетики поведения. На мышах разных линий, как инбредных, так и селектированных, исследованы генетические вариации в поведении и корреляция иногда достаточно сложных признаков поведения с изменчивостью строения некоторых отделов мозга. Мыши широко используются также для изучения нейро-биологических основ процесса обучения, причем все большую роль начинают играть исследования поведения и способности к обучению у мышей, у которых генноинженерными методами определенные гены либо выключены (мыши-нокауты), либо видоизменены (искусственные мутанты). Изучение таких животных методами генетики поведения дает также возможность моделировать целый ряд неврологических и психических заболеваний человека (эпилепсия, алкоголизм, депрессивные состояния, болезнь Алыдгеймера и др. — Driscoll, 1992).