Асимметрия мембран
Однако имеются мембраны, в которых миграция липидов протекает очень быстро, с порядка нескольких минут. Такие данные получены для эндоплазматического ретикулума печени крысы, а также для цитоплазматической мембраны грамположитель-ных бактерий В. megaterium. В этих мембранах происходит синтез липидов, и в них, по-видимому, присутствуют специальные транслоказы, которые обеспечивают быструю трансмембранную миграцию липидных молекул. Такое предположение было высказано в отношении эндоплазматического ретикулума, но оно пока не нашло экспериментального подтверждения. Характерное время трансмембранной миграции липидов в мембране эритроцитов имеет промежуточное значение и составляет величины порядка нескольких часов в зависимости от структуры изучаемого липида. Такие же результаты были получены при измерении скорости флип-флопа спин-меченных аналогов фосфолипидов и экзогенных лизо-фосфолипидов. Было установлено, что скорость миграции возрастает при нарушениях цитоскелета, а также под действием агентов, влияющих на структуру липидиого бислоя. Возможно, цитоскелет играет определенную роль в уменьшении скорости миграции липидов через бислой благодаря связыванию аминофосфолипидов. Характерно, что ни эндоплазматический ретикулум, ни бактериальная ци-топлазматическая мембрана, для которых характерна высокая скорость флип-флопа липидов, не связаны с цитоскелетом.
Тот факт, что скорость трансмембранной миграции липидов в эритроцитах является АТР-зависимой, предполагает присутствие в этой мембране энергозависимой транслоказы. АТР-зависимая трансмембранная миграция аминофосфолипидов наблюдается также в плазматической мембране фибробластов и лимфоцитов. Однако ни одна из фосфолипидных транслоказ до сих пор не выделена, поэтому возможность существования таких ферментов и их потенциальная роль в поддержании липидной асимметрии или в биогенезе мембран представляются вероятными, но не более того.
Латеральная гетерогенность мембран
Исходная жидкостно-мозаичная модель предполагает, что распределение белковых и липидных компонентов в плоскости бислоя является гомогенным. Однако не вызывает сомнений, что в ряде мембран существуют домены или области, отличающиеся по составу от остальной части мембраны вследствие ограничений в диффузионном обмене их компонентов. Имеются различные виды мембранных доменов, о которых можно говорить в рамках жидкост-но-мозаичной модели, вводя определенные ограничения, налагаемые дополнительными стабилизирующими эти домены взаимодействиями.
1.Макроскопические домены, как правило, представляют собой обширные участки на поверхности клетки с характерной морфологией и четкими границами. Примерами являются апикальная и базолатеральиая области поляризованных эпителиальных клеток. В тилакоидах соприкасающиеся и несоприкасающиеся участки фото-синтезируюших мембран тоже имеют разный состав и, по-видимому, стабилизируются межмембранными взаимодействиями в стопках.
2.Агрегация белков в плоскости мембраны может приводить к образованию довольно больших островков, или доменов, которые обогащены определенным белком и находятся в смеси с какими-либо другими компонентами. Примерами являются пурпурные мембраны Н. halobium, содержащие бактериородопсин, или щелевые контакты, содержащие коннексин.
3.Домены, формируемые при участии цитоскелета, в принципе могут образоваться путем ассоциации определенных мембранных белков за счет их взаимодействия с внутриклеточными белками. В основе такой латеральной организации мембранных белков могли бы лежать особенности взаимодействий, наблюдаемых в случае цитоскелета эритроцитов. Пока четкие примеры существования таких доменов отсутствуют, однако можно предположить, что пэтчинг и кэппинг антигенов на клеточной поверхности осуществляются именно с участием цитоскелета и что концентрирование специфичных рецепторов в окаймленных ямках плазматической мембраны перед эн-доцитозом осуществляется благодаря их взаимодействию с компонентами цитоскелета или с клатрином.
4.Липидные микродомены могут быть термодинамически стабильны как в биологических мембранах, так и в модельных липид-ных системах. Об этом свидетельствуют многочисленные косвенные данные, хотя четкие доказательства существования таких доменов пока отсутствуют.
Приведенная классификация доменов условна; указанные четыре категории отнюдь не исключают друг друга при рассмотрении способов стабилизации латеральной гетерогенности мембран.
МАКРОСКОПИЧЕСКИЕ ДОМЕНЫ И БАРЬЕРЫ В ПЛАЗМАТИЧЕСКОЙ МЕМБРАНЕ
Плазматическая мембрана клеток часто бывает разделена на отдельные домены, которые можно даже выделить и охарактеризо-

вать. Как правило, эти домены разделены барьерами, которые препятствуют переходу белков и, возможно, липидов из одного домена в другой. В пределах же областей, ограниченных этими барьерами, белки и липиды диффундируют свободно. На рис. 4.7 приведено несколько примеров таких доменов.
1.Апикальная и базолатеральная области мембраны поляризованных эпителиальных клеток имеют разный состав. Показано, например, что ганглиозиды не пересекают границу между этими областями, которая в данном случае представляет собой область плотных контактов между клетками.
2.Плазматическая мембрана спермиев состоит из четко разграниченных участков разного состава, которые можно разделить. Граница между доменами препятствует свободной диффузии мембранных белков.
3.Соединительный отросток на палочке сетчатки разделяет ее наружный и внутренний сегменты. Родопсин исходно включается в мембрану внутреннего сегмента, а затем концентрируется в мембране наружного сегмента. Вероятно, в мембране палочки существует диффузионный барьер, поддерживающий эту разность концентраций.
4.Натриевые и калиевые каналы локализованы в разных областях мембраны миелинизированного аксона. Возможно, такая организация стабилизируется благодаря контактированию мембраны с глиальиыми или шванновскими клетками, которые образуют миелиновую оболочку вокруг нервного волокна.
Следует отметить, что макроскопическая латеральная гетерогенность может быть характерна и для мембран прокариотических клеток. Например, в мембранах грамотрицательных бактерий имеются области адгезии, в которых, по-видимому, осуществляется контакт между наружной и внутренней мембранами. У пурпурных несерных фотосинтезирующих бактерий фотосинтетический аппарат локализован в специализированных мембранах, образующихся в результате инвагинаций цитоплазматической мембраны.
Экспериментальные исследования в этой области затруднены, поэтому о природе барьеров, разделяющих различные мембранные домены, известно очень мало.
ТИЛАКОИДНЫЕ МЕМБРАНЫ
Мембраны тилакоидов в хлоропластах высших растений содержат фотосинтетический аппарат. Эти мембраны собраны в стопки, называемые гранами. Соприкасающиеся и несоприкасаю-
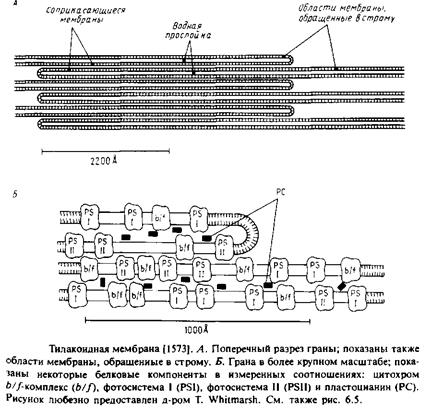
щиеся участки мембраны тилакоидов имеют разную морфологию, и их можно разделить. Известно также, что эти две области мембраны имеют разный состав, вероятно стабилизируемый благодаря взаимодействиям между мембранами. Одним из факторов стабилизации мембран в стопках является прямое связывание светособирающих комплексов друг с другом в соприкасающихся мембранах. Важную роль в стабилизации стопок могут играть и электростатические взаимодействия. Детали этих взаимодействий во многом неясны, однако очевидно, что они каким-то образом приводят к функционально значимому латеральному разделению компонентов. Например, две фотосистемы, I и II, которые являются компонентами электронтранс-портной цепи, расположены в разных мембранных доменах, но связаны биохимически с помощью диффундирующего пластохинона. Распределение светособирающего комплекса между этими двумя доменами зависит от степени его фосфорилирования.