Страница
6
ПРИМЕРЫ ЛИПИДНОЙ АСИММЕТРИИ
Из-за экспериментальных трудностей, связанных с определением липидной асимметрии, можно привести лишь несколько примеров, в которых асимметрия несомненно доказана. Прежде всего следует упомянуть эритроциты человека, которые в этом отношении были, детально изучены, причем все использованные методы дали хорошо согласующиеся между собой результаты. Другой пример такого рода — это высокая степень асимметрии наружной мембраны грамот-рицательных бактерий; впрочем, эта мембрана необычна в том отношении, что ее основным компонентом является уникальный липо-полисахарид. Нет никаких сомнений, что асимметричное распределение липидов свойственно и другим биологическим мембранам, однако убедительные данные получены лишь в немногих случаях.
Липидная асимметрия в фосфолипидиых везикулах
Для малых моноламеллярных везикул, состоящих из двух разных липидов, характерно асимметричное распределение липидов. Например, в везикулах, состоящих из смеси фосфатидилхолина с другими липидами, наружный монослой обогащен сфингомиелином или фосфатидилглицеролом, а фосфати-дилсерин, фосфатидилэтаноламин, фосфатидилинозитол и фосфа-тидная кислота предпочитают находиться на внутренней поверхности. Общепризнано, что в этих случаях липидная асимметрия возникает прежде всего из-за различий в упаковке молекул на двух сторонах бислоя ММВ. Липиды с более объемными полярными головками стремятся находиться в наружном монослое, потому что там больше площадь поверхности, приходящейся на молекулу. Однако биологические мембраны, за отдельными исключениями, не имеют участков со столь большой кривизной, так что эти выводы нельзя безоговорочно распространить на любые биологические мембраны. С помощью трансмембранного градиента рН липидную асимметрию можно индуцировать и в больших моноламеллярных везикулах.
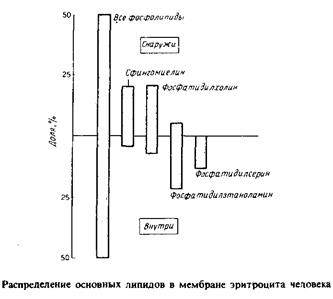
Липидная асимметрия в эритроцитах человека
Четко показано, что распределение липидов в мембране эритроцитов в высшей степени асимметрично. Как видно из рис. 4.4, фосфа-тидилхолин и сфингомиелин находятся преимущественно в наружном монослое, тогда как фосфатидилэтаноламин и фосфатидилсе-рин — в основном во внутреннем.
Имеются убедительные данные о том, что для сохранения липидной асимметрии необходима целостность цитоскелета. Она нарушена в клетках, дефицитных по спектрину или белку полосы 4.1. Воздействия, нарушающие цитоскелет, изменяют и липидную асимметрию, хотя неясно, являются ли эти эффекты прямым следствием повреждения цитоскелета. Кроме того, в малых везикулах, полученных из мембраны эритроцитов, распределение липидов менее асимметрично. Эти данные показывают, что цитоскелет играет определенную роль в поддержании липидной асимметрии. Механизм такого влияния неясен, однако можно предположить, что имеет место прямое связывание цитоске-летных белков с аминофосфолипидами.
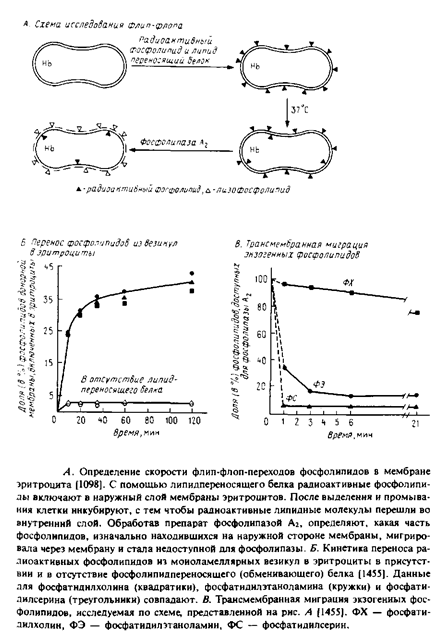
Прямая стабилизация асимметрии, если она существует, — это только одна сторона вопроса. Так, экзогенные фосфолипиды, включенные в мембрану эритроцитов с помощью обменивающих белков, примерно через сутки перераспределяются, и их асимметрия принимает такой же характер, как и у эндогенных липидов. На рис. 4.5 приведена схема этих экспериментов. Измеряя кинетику гидролиза под действием фосфолипаз, можно определить скорость флип-флопа липидных молекул. Эта скорость возрастает в ряду фосфатидилсерин > фосфатидилэтаноламин > фосфатидил-холин > сфиигомиелии. Подобные результаты были получены с помощью спин-мечеиных липидов и лизофосфолипидов. Таким образом, те липиды, которые локализуются на внутренней стороне мембраны, имеют и относительно большую скорость транс-мембраииой миграции. Те же липиды, которые находятся на наружной стороне, мигрируют значительно медленнее либо вообще не совершают флип-флоп-перескоков.
Очень важным является вывод о том, что быстрая трансмембранная миграция аминофосфолипидов, по-видимому, является АТР-зависимой и значительно замедляется в клетках, дефицитных по АТР. Это послужило основанием для предположения о том, что специфичный флип-флоп липидов катализируется особыми ферментами типа транслоказ. В пользу этого предположения появляется все больше данных, правда, многие из них являются косвенными.
Есть два взгляда на то, как поддерживается липидная асимметрия в мембранах; они взаимно дополняют друг друга и могут быть в равной степени важны. Один из них дает статическую картину с акцентом на стабилизацию асимметрии за счет специфических взаимодействий фосфолипидов с цитоскелетными белками, а другой представляет асимметрию как динамический феномен, когда энергозависимые транслоказы избирательно переносят липиды через бислой, поддерживая их стационарное асимметричное трансмембранное распределение.
Липидная асимметрия наружной мембраны бактериальных клеток
Наружная мембрана грамотрицательных бактерий в отличие от мембраны эритроцитов не может служить моделью других бактериальных мембран. Это связано с тем, что содержащийся в ней липо-полисахарид является в своем роде уникальным мембранным компонентом. Детальные исследования показали, что эта мембрана представляет собой высокоасимметричную структуру. Указанный липополисахарид был обнаружен только на наружной стороне бислоя, а большая часть фосфолипидов локализована во внутреннем монослое, обращенном в периплазму. Липополисахарид играет важную роль как барьер для проникновения внутрь клетки некоторых веществ; в частности, именно благодаря ему бактерии приобретают устойчивость к ряду антибиотиков. Одним из основных компонентов наружной мембраны явлется так называемый липопротеин Брауна, который закреплен в мембране с помощью ковалентно связанных с ним липидов, а кроме того, связан с пептидогликановой стенкой за счет ковалентных и нековалентных взаимодействий.
ТРАНСМЕМБРАННАЯ МИГРАЦИЯ ЛИПИДОВ
Биосинтез фосфолипидов и сборка мембраны протекают асимметрично. Активные центры ферментов биосинтеза фосфолипидов локализованы на одной, а не на двух сторонах мембраны. Например, фосфолипиды синтезируются и внедряются в мембрану на цитоплазматической стороне эндоплазматического ре-тикулума печени крысы и на внутренней стороне бактериальной цитоплазматической мембраны. Ясно, что эти липиды должны пересечь мембрану, чтобы достичь противоположной стороны бислоя.
Скорость трансмембранной миграции фосфолипидов в фосфоли-пидных везикулах пренебрежимо мала: ее характерное время составляет несколько суток. Флип-флоп-переход может ускоряться в присутствии таких интегральных мембранных белков, как гликофорин, или при возмущениях в бислое, происходящих, например, при обработке фосфолипазами. Как и следует ожидать, перемещение липидных молекул затрудняют именно полярные головки, поскольку производные диацилглицерола очень быстро мигрируют через бислой. Для некоторых биологических мембран, например мембраны вируса гриппа и внутренней мембраны митохондрий, также характерна очень малая скорость трансмембранной миграции фосфолипидов.