Страница
4
Предполагается, что именно благодаря взаимодействиям цитоскелета с мембраной возникает трансмембранное распределение ли-пидов, стабилизируются латериальные белковые домены и обеспечивается направленное перемещение белков в мембране.
Цитоскелетную сеть образуют три типа волокон: 1) микрофила-менты, состоящие из актина и связанных с ним белков; 2) промежуточные филаменты, состоящие из кератинов и родственных им белков; 3) микротрубочки, состоящие из тубулина. В биохимическом отношении лучше всего изучено связывание с мембраной актиновых мик-рофиламентов. Особого рассмотрения здесь заслуживает актин-спектриновая сеть эритроцитов. Эти белки способствуют объединению актиновых волокон в пучки, прикрепляют филаменты к мембране, образуют сшивки в актине, регулируют длину филаментов, влияют на их сократительную способность и обеспечивают отклик на Са2 +. В ряде случаев актин участвует в сократительной активности. Микрофиламенты разрушаются цитоха-лазинами.
Микрофиламенты могут располагаться параллельно цитоплазматической мембране, как в сократимом кольце в делящейся клетке, или могут быть связаны с плазматической мембраной одним своим коццом, как в местах адгезии в области контакта клетка—клетка или клетка—субстрат. Полагают, что связывание актина с мембранами обеспечивается несколькими мембранными белками. Из данных о физической близости микро-филаментов к мембране, о биохимических взаимосвязях микрофила-ментов и разрушающем воздействии цитохалазина следует, что микрофиламенты участвуют во многих мембранных процессах, в частности в опосредуемом рецепторами эндоцитозе, пэтчинге и кэппинге, клеточной подвижности и цитокинезе. С биохимической точки зрения лучше всего охарактеризованы взаимодействия актина с мембраной у эритроцитов.
ПРОМЕЖУТОЧНЫЕ ФИЛАМЕНТЫ
Это полимеры, состоящие из одного или двух фибриллярных полипептидов, которые различаются в клетках разного типа и кодируются семейством мультигенов. Примером являются кератины из эпителиальных клеток и виментин из клеток мезенхимы.
Функции промежуточных филаментов неизвестны. Мало что можно сказать и о биохимической основе их взаимодействия с мембранами.
МИКРОТРУБОЧКИ
Они состоят из тубулина, который хорошо охарактеризован и представляет собой а/?-гетеродимерный белок. Микротрубочки образуют цитоплазматическую сеть, которая, как полагают, связывает плазматическую мембрану с органеллами, например с митохондриями. Вдоль микротрубочек, по-видимому, происходит перемещение эндосом и лизосом. Есть доказательства, что тубулин прикрепляется к мембранам в особых точках. Выделен мембранный белок синапсин I, который, по-видимому, взаимодействует с тубулином. В ходе митоза цитоплазматическая сеть микротрубочек распадается и перестраивается в митотическое веретено. Микротрубочки разрушаются под действием колхицина.
МЕМБРАНА И ЦИТОСКЕЛЕТ ЭРИТРОЦИТОВ
Наиболее детально изучены мембрана и цитоскелет эритроцитов млекопитающих. В табл. 4.2 перечислены основные белки, которые были разделены с помощью ДСН-ПААГ-электрофореза. Цифровые обозначения полипептидов связаны с их относительной электрофоретической подвижностью в геле. При промывании мембраны растворами с низкой ионной силой удаляются периферические мембранные белки, к которым прежде всего относятся компоненты цитоскелета. Основными интегральными белками цитоскелета являются белок полосы 3 и гликофорины А, В и С. Белок полосы 3 представляет собой анионный переносчик, а функции гли-кофоринов, относящихся к классу гликопротеинов, неизвестны. В электронном микроскопе цитоскелет выглядит как упорядоченная сеть на внутренней стороне мембраны. Как видно из табл. 4.2, белки цитоскелета являются основными мембранными компонентами, и это облегчает их биохимическую характеристику. По сути белковый каркас состоит из спектрин-актинового комплекса, который связан с плазматической мембраной благодаря взаимодействиям как с белком полосы 3, так и с гликофорином; эти взаимодействия осуществляются с помощью специальных белков — ан-
Таблица 2. Свойства, степень ассоциации и функции эритроцитарных мембранных белков

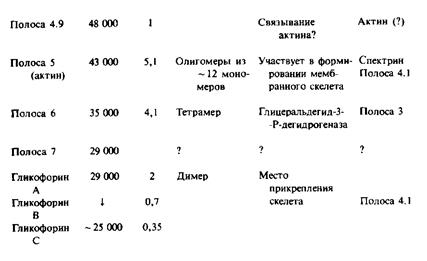
кирина и белка полосы 4.1. Основные компоненты были очищены до гомогенного состояния и изучены in vitro. Комплексы между основными белками, такими, как белок полосы 3 и анкирин или анкирин и спектрин, характеризуются константами диссоциации порядка Ю-7 М, которые могут меняться в физиологических условиях. Фосфорилирование анкирина влияет на его аффинность по отношению к спектрину, а взаимодействие между белком полосы 4.1 и гликофорином, по-видимому, модулируется фосфатидилинози-толами.
Интересно, что белки, близкородственные компонентам цитоскелета эритроцитов, обнаружены в ряде неэритроидных клеток. Большой интерес к цитоскелету эритроцитов, по всей вероятности, обусловлен тем, что данная система не является уникальной лишь для этих клеток, а представлена в виде кортикального цитоскелета и в клетках другого типа. Рассмотрим свойства некоторых цитоскелет-иых белков.
1. Спектрин. Это тетрамер типа г, в котором два гетероди-мера a/J объединены по схеме «конец-к-концу». Молекула может достигать в длину 2000 А. Спектрин связан с анкирииом и белком полосы 3 по сайтам, расположенным на противоположных концах молекулы. Кроме того, спектрин связан с актином, возможно, в комплексе с белком полосы 4.1. Спектринопо-

добные молекулы, например фодрин, обнаружены в клетках разного типа.
2.Актин. Это глобулярный белок, который существует в виде линейных олигомеров, содержащих по 12—18 молекул. Они выглядят на электронных микрофотографиях как короткие стержни, к которым может быть прикреплено до шести спектриновых тетрамеров.
3.Анкирин. Это наиболее охарактеризованный растворимый белок, обеспечивающий взаимодействия между интегральными мембранными белками и цитоскелетом. Он имеет отдельные домены, ответственные за независимое связывание со спектрином и с цитоп-лазматическим доменом белка полосы 3. Анкирин был обнаружен и в неэритроидных клетках.
4.Белок полосы 4.1. Он также относится к классу белков, обеспечивающих связь цитоскелета с мембраной. Белок полосы 4.1 связывается со спектрином и актином, а также с гликофорином. Кроме того, при определенных условиях он может связываться и с белком полосы 3. Белку полосы 4.1, по-видимому, родствен синапсин I, обнаруженный в мембране синаптических везикул.
5. Белок полосы 3. Это основной анионный переносчик в эритроцитах. Цитоплазматический домен содержит на N-конце кислый участок, который связывается с некоторыми гликоли-тическими ферментами, а также с гемоглобином. Цитоплазматический домен не участвует в транспорте анионов. Его сегмент, расположенный вблизи мембраны, связывается с анкири-ном и с белком полосы 4.2. На основании анализа аминокислотной последовательности было высказано предположение, что белок полосы 3 имеет 12 трансмембранных сегментов, но полученные к настоящему времени экспериментальные данные не позволяют ни подтвердить, ни опровергнуть это положение.