Кальций – фосфорно – магниевый метаболизм
Процессы депонирования кальция и фосфора в костях и их абсорбции/экскреции в кишечнике и почках взаимно сбалансированы так, что концентрация этих ионов в крови изменяется в весьма узких пределах (8,8-10,4 мг/дл или 22-26 мМ/л кальция и 2,5-4,5 мг/дл или 9-13 мМ/л фосфата).
Магний — преимущественно, внутриклеточный катион, четвертый по абсолютному содержанию в организме (Л.Г.Смит, 1987). Тело взрослого человека содержит около 25 г магния. В интрацеллюлярной жидкости его концентрация в 8 раз выше, чем в интерстициальной. Взрослому человеку в день нужно не менее 3,5—4,5 мг магния, чтобы не расходовать его костные резервы. Богаты магнием зелень, где он выполняет ключевую роль при фотосинтезе в составе хлорофилла, морепродукты и мясо, орехи и семечки, бобовые, бананы и цитрусовые, шоколад, патока и кокосы. Впрочем, если этих разносолов на столе нет, полезно помнить, что очень богаты данным металлом маковые зерна, а также самый обыкновенный . чай.
Магний всасывается в тонком кишечнике, при участии витамина D, примерно на 40% от его поступления с пищей. Избыток фитиновой кислоты и жирных кислот, а также алкоголь отрицательно влияют на его абсорбцию. Высокие концентрации магния в кишечном содержимом мешают всасыванию кальция, но не наоборот. Магний экскретируется почками, причём эффективность его реабсорбции может достигать 95%. Почки варьируют экскрецию магния в равновесном, по отношению к поступлению этого электролита, режиме, в широчайшем диапазоне — от 1 до 250 мМ в день. Алкоголь препятствует реабсорбции магния в нефронах. Кальций и магний конкурируют при реабсорбции. Магний — составная часть минерального вещества костей, участник работы трансфосфорилирующих ферментов и амино-ацил-т-РНК-синтетаз, обеспечивающих условия для трансляции белков. В электрофизиологических процессах определённое значение имеет роль магния, как антагониста кальция, проявляющаяся в их различном влиянии на ЦНС.
Центральные органы, регулирующие кальций-фосфорно-магниевый обмен — паратиреоидные железы.
В гистологии данные органы называются околощитовидными или паратиреоидными.
Нижние паращитовидные железы возникают из того же третьего глоточного кармана, который дают начало и тимусу (см. выше), а верхние являются дериватами четвёртого глоточного кармана. Таким образом, у верхних и нижних полюсов каждой из долей щитовидной железы, вне капсулы последней, но под её фасцией, в норме, обнаруживается по одной паращитовидной железе.
Однако, топография данного эндокринного органа, быть может, наиболее изменчива среди всех эндокринных желез. У очень значительной части людей (более 10%) дополнительные паращитовидные тельца обнаруживаются по всему ходу эмбриональной миграции глоточных карманов: в том числе, в тимусе, переднем средостении, близ каротидных артерий. Они служат нередко источником эктопических гормонообразующих опухолей.
Паращитовидные железы — наиболее «молодое» органное открытие эндокринологов. Верхние наружные паращитовидные железы впервые описал шведский анатом И.К. Сундстрём, только в 1880 г.
Паращитовидные железы состоят из капсулы, стромы и недольчатой паренхимы, в которой представлены мелкие главные клетки двух подвидов: тёмные, содержащие секреторные гранулы и, вероятно, покоящиеся; и светлые — лишённые таких гранул и секреторно активные, последние также богаты гликогеном. В железе имеются также более крупные оксифильные клетки, появляющиеся, очевидно, путём трансформации главных в период пубертата и с возрастом всё более многочисленные. Оксифильные клетки рассматриваются как результат инволюции главных. Функция оксифильных клеток точно не известна доныне. По последним данным, парат-гормон может синтезироваться и в них.
Главные клетки обладают очень развитым гладким эндоплазматическим ретикулюмом (ГЭР), в оксифильных ГЭР представлен слабее. Оксифильные клетки богаты, а главные — относительно бедны митохондриями. Интересная особенность нормальной структуры паращитовидных желез — наличие в каждой из них большого количества жира, накапливаемого с возрастом (у пожилых — до 70% объема желез). При гиперплазии и опухолях количество жира в паращитовидных железах резко снижается.
Основной продукт паращитовидной железы — паратиреокринин (прежние названия — паратирин или парат-гормон). Структуру паратироидного гормона расшифровали в 1970 г. Х.Д.Найел и соавторы. Его выделяют главные клетки.
Парат-гормон — это пептидный регулятор, (рис. 2), состоящий из 84 аминокислот (молекулярной массой чуть более 9,5 кД).
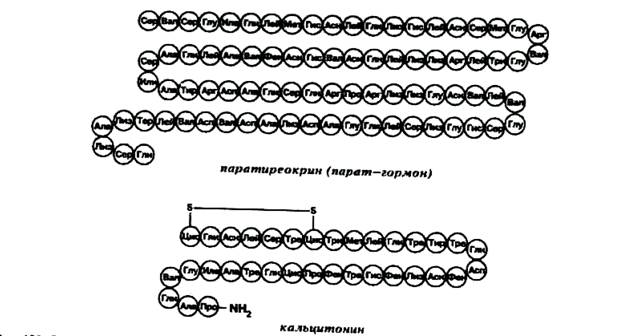
Рис. 2. Структура парат-гормона и кальцитонина человека.
Парат-гормон возникает из препрогормона длиной в 131 аминокислотный остаток (молекулярной массой около 12,5 кД, синтезируется на полисомах), через стадию прогормона (90 аминокислот, образуется в ЭПР под действием клипазы), причем его процессинг модулируется ферментом фурином. Прогормон поступает за счет энергозависимого механизма в комплекс Гольджи, где протеолитический мембранно-связанный комплекс (триптическая клипаза) вычленяет из него активный гормон. Препрогормон кодируется в 11 -и хромосоме, а фурин — в 15-й. Оба экспрессируются совместно. Весь процесс синтеза и секреции (которая может происходить как в виде экзоцитоза специальных гранул, так и в безгранулярном режиме.) занимает около 30 мин., причём 15 мин. тратится на упаковку готового гормона в гранулы.
Паратиреоидная секреция активируется, в основном, в ответ на снижение концентрации ионизированного (свободного) кальцияв крови. Опосредованно, гиперфосфатемия также активирует паращитовидные железы, снижая концентрацию ионизированного кальция. Также, как кальций, но значительно слабее, на секрецию парат-гормона влияет и магний. Однако тяжелая длительная гипомагниемия парадоксальным образом подавляет секрецию парат-гормона, так как магний необходим самим паратиреоцитам для выделения их гормонов (см. ниже). Главные клетки располагают кальциевым сенсором — трансмембранным гликопротеином, вмонтированным в их плазматическую мембрану. Таким же сенсором обладают, кроме паратироцитов, С-клетки щитовидной железы и некоторые клетки мозга и почек. Этот рецептор кодируется в хромосоме 3, при повышении уровня экстрацеллюлярного ионизированного кальция он блокирует экспрессию генов гормона паращитовидных желез и ключевого фермента его активации. В настоящее время доказано, что выработка парат-гормона, преимущественно, регулируется in vivo на посттранскрипционном уровне. При повышении уровня иона кальция в крови происходит стимуляция рецептора, активация пострецепторного Сq-белка и нарастание концентрации кальция в цитозоле, блокирующее функцию главных клеток. Мутации данного сенсора дают при гомозиготности тяжелый наследственный неонатальный гиперпаратиреоз, а у гетерозигот — доброкачественную семейную гипо-кальциурическую гиперкальциемию (см. ниже).