Страница
3
В нижней, подземной, части стебля находится корневище стелящееся по почве, у бриевых мхов имеются нити из лишенных хлорофилла клеток – ризоиды, которые выполняют функцию закрепления на субстрате и проведения воды и минеральных солей к стеблю (рис 13). Ризоиды образуются на ранней стадии прорастания споры, параллельно с протонемой. Протонема может быть нитчатой (похожей на водоросли) и пластинчатой (рис 14). Протонема с почками возобновления листостебельных растений (поздняя стадия развития протонемы) называется каулонемой (рис 15).
|
Рис 13. Гаметофит мха с ризоидами |
Рис 14. Пластинчатая (1) и нитчатая (2) протонемы мхов |
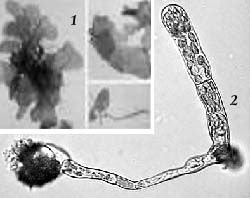
Рис 15. Каулонема (протонема с почками)
Анатомия
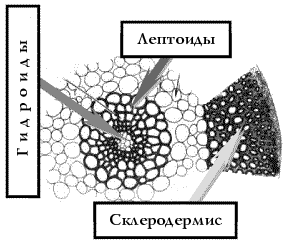
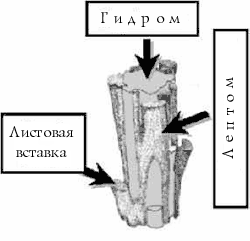
Стебель имеет относительно сложное строение (рис 16, 17). Внешний слой образует эпидермис, из клеток которого могут вырастать многоклеточные нити – ризоиды. Следующий за эпидермисом слой называется склеродерма. Он состоит из узких и длинных толстостенных клеток, выполняющих проводящую и механическую функции. Самый внутренний слой клеток называется центральным пучком, он состоит из сосудов, очень напоминающих сосуды других высших растений. Различают два типа клеток сосудов – лептоиды (похожие на клетки флоэмы) и гидроиды (подобные клеткам ксилемы). Совокупность лептоидов в стебле называют лептом, а гидроидов – гидром (рис 18). Помимо лептоидов и гидроидов в центральном пучке имеются стереиды, сходные с волокнами склеренхимы сосудистых растений. В месте прикрепления стеблевого листа к стеблю от центрального цилиндра ответвляется пучок лептоидов и гидроидов, образующих листовую вставку (аналогичную листовому следу у сосудистых). Анатомия стебля сильно варьирует у разных представителей бриофитов, у конкретных видов в состав стебля могут входить не все перечисленные компоненты или они могут по-разному комбинироваться.
|
Рис 16. Схематическое строение стебля Bryales |
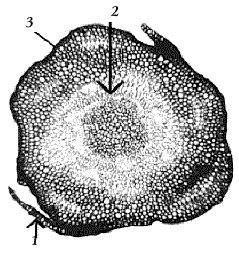
Рис 17. Поперечный срез стебля кукушкина льна: 1 – лист; 2 – центральный пучок; 3 – эпидермис |
|
Рис 18. Трехмерная диаграмма строения стебля мха |
Рис 19. Транспорт воды гидроидами: 1 – центральный пучок; 2 – листовая вставка |
Если сравнивать строение ножки спорофита со стеблем, можно обнаружить, что в состав и того и другого входят гидроиды. Анатомия гидроидов очень сходна с анатомией трахеид сосудистых: их клетки имеют толстые стенки и удлиненные острые окончания. Однако у гидроидов, в отличие от трахеид, отсутствуют спиральные целлюлозные волокна, а их стенки лишены лигнина. Гидроиды осуществляют проводящую функцию (рис 19) Строение лептоидов сходно со строением ситовидных трубок. Листья Polytrichum также имеют сложное строение. Линейно-ланцетная пластинка (рис 20) их почти на всем протяжении многослойна и состоит, в общем, из тех же элементов, что и стебель. На верхней стороне листа находятся особые ассимиляторы. Это довольно высокие пластинки, идущие вдоль листа на близком друг от друга расстоянии. Ассимиляторы несут работу по ассимиляции углерода. Строение листьев обусловливают поверхностное накопление влаги и приводят к заболачиванию.
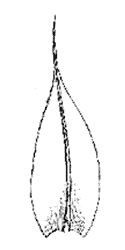
Рис 20. Лист Bryales с центральной жилкой
Жизненный цикл
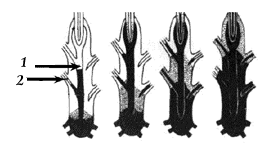
Из гаплоидной споры вырастает первичный росток – протонема (рис 14), которая развивается в протонему с почками – каулонему (рис 17). Со временем из почки вырастает тоже гаплоидное листостебельное растение – гаметофор. Вначале оно молодое, недоразвитое, но вскоре набирает силы и приобретает привычный облик. Совокупность гаметофора, протонемы и каулонемы называется гаметофитом, или гаплоидным поколением мха. У гаметофора мхов есть одна примечательная особенность. Выражаясь образным языком, он имеет вечно живущую часть – точку роста (рис 22), срок жизни которой не ограничен (даже зимой), и временно живущую часть, или «медленно разлагающийся труп», – все остальное листостебельное растение, срок жизни которого определен сезоном вегетации. На гаметофоре со временем образуются гаметангии – антеридии – ♂ (рис 23, 1) и архегонии ♀(рис 23, 2).Они располагаются на вершине стебля целыми собраниями, отдельно друг от друга. Собрания антеридиев, или, как их иногда называют, «мужские цветки», имеют вид розеток, окруженных широкими желтоватыми или красноватыми листьями. Центр их занят антеридиями. Антеридий представляет собой «мешочки на ножках» - удлиненный мешок, сидящий на короткой ножке со стенкой из одного слоя клеток. Внутри антеридиев созревают антерозоиды – мужские половые клетки, способные к самостоятельному передвижению в водной среде. Антеридии вырастают в пазухах веточных листьев, которые отличаются по своему строению от стерильных, имеют красноватую окраску и называются антеридиальными (рис 23, 2). Совокупность антеридиев и покрывающих их листьев на веточке называют перигонием. Архегонии вырастают на стебле, как боковые веточки. Сверху они прикрыты перихециальными листьями (рис 23, 1). Совокупность архегония и покрывающих его листьев называют перихецием. Архегонии устроены более сложно (рис 23, 2). По форме они напоминают колбочку. В нижней (широкой) части архегонии имеют двухслойную стенку. Эта его часть называется брюшком. Узкая часть архегония – «горлышко колбочки» – имеет одноклеточную стенку. Эта часть называется шейкой. Она сверху закрыта легко опадающим колпачком с тонкими, направленными вниз волосками, напоминающими льняную пряжу («кукушкин лен»). В брюшке помещаются яйцеклетка и брюшная канальцевая клетка, в шейке – длинный ряд шейковых канальцевых клеток. Сидит архегоний на массивной многоклеточной ножке. Антеридии и архегонии у видов сырых местообитаний образуются на гаметофоре ранней весной, сразу после схода снега, пока талая вода, необходимая для передвижения антерозоидов, не сошла. Под влиянием воды, попадающей в «мужской цветок», антеридии вскрываются на вершине, и сперматозоиды выходят в воду. В то же время вскрываются на верхушке и архегонии; при этом все канальцевые клетки расплываются в слизь, заполняющую канал архегония. Сперматозоид, представляющий собою спирально изогнутое и снабженное двумя жгутиками тельце, проникает через эту слизь в архегоний; достигнув яйцеклетки, он сливается с нею и производит оплодотворение. После опоражнивания антеридиев стебель, на котором помещался «Мужской цветок», продолжает свой рост. Оплодотворенная же яйцеклетка начинает делиться и дает начало спорофиту – спороносному поколению мха с диплоидным набором клеток.
|
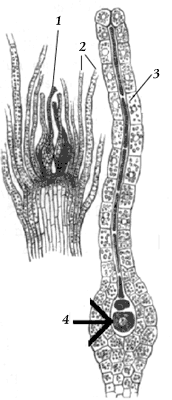
Рис 22. Апикальная точка роста мха (показана стрелкой) |
Рис 24. Архегоний и перихеций: 1 – архегоний; 2 – перихециальные листья; 3 – шейка архегония; 4 – яйцеклетка |
|
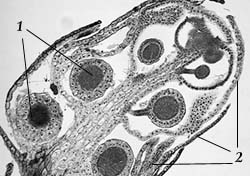
Рис 23. Антеридиальная веточка мха: 1 – антеридии; 2 – антеридиальные листья |