Что такое молекулярный ламаркизм
В самом деле, Вейсман сам был вынужден отказаться от крайних своих утверждений. В 1892 г. он писал: «Корень наследственных изменений должен лежать глубже [чем в слиянии родительских половых клеток. – Ю.Ч.] и заключаться в прямом воздействии внешних влияний на биофоры [единицы наследственности – Ю.Ч.]». В чем же состоит такое воздействие? Этого, разумеется, Вейсман не знал и, в стиле натурфилософов прошлого, написал: «Начало изменения основывается на малых неправильностях в питании зародышевой плазмы». Тут-то Стил и смог дать радикальное уточнение, невозможное в дни Вейсмана: «питание» наследственного материала состоит в усвоении им генетического материала некоторых вирусов.
Конечно, сводить эволюцию, даже только приспособительную, к одному лишь наследованию приобретенных признаков наивно, зато такое сужение задачи позволило Стилу сразу вчерне решить ее. По его мнению, зародышевая плазма действительно играет ведущую роль в эволюции, поскольку ее фрагменты передаются между органами и организмами с помощью вирусов. Тем самым, вирусы, по крайней мере некоторые их формы, например ретровирусы (вирусы, РНК которых передает свою информацию в хромосому организма путем обратной транскрипции), являются не только и даже не столько причиной болезней, сколько необходимым эволюционным агентом.
Главная мысль новой книги Стила с соавторами проста и убедительна: синтез антител есть создание новой генетической информации, не сводимое к случайным ненаправленным вариациям «по Дарвину», причем нужный ген формируется целенаправленно. Тем самым феномен приобретенного пожизненного иммунитета выступает как несомненный факт наследования приобретенного признака, причем наследование происходит путем включения вновь найденного гена, кодирующего антитело, в хромосомы клеток иммунной памяти, из которых при надобности вновь производятся клетки, способные производить антитела.
Исходя из этого факта делается правдоподобное допущение: эволюционно значимая информация может передаваться из хромосом стволовых клеток лимфоидной системы в хромосомы стволовых клеток половой железы. А осуществляют эту передачу, по Стилу, ретровирусы. Это допущение еще предстоит проверить. Пока же поговорим о механизме приобретения новой генетической информации в ходе иммуногенеза и подумаем, что это дает для понимания эволюции.
5. Генетический принцип обеспечения разнообразия антител
У зародыша млекопитающих есть совсем немного генов, кодирующих иммуноглобулины, – около сотни. Их последующее разнообразие в ходе развития организма каждый раз создается заново, точно так же, как заново создается любой орган. Создается оно путем комбинирования фрагментов существующих генов. Но этого разнообразия оказывается мало, поэтому конкретное антитело обычно не выбирается из наличных, а создается в ответ на конкретную заразу (на антиген). Процесс идет так.
В стрессовой ситуации, которую создает вторжение антигена, включается механизм перестройки генов иммуноглобулинов: генетическая система по каким-то не вполне еще понятным правилам режет и сшивает фрагменты генов до тех пор, пока не найдет приемлемый вариант – тот, что синтезирует антитело, которое реагирует с вторгшимся антигеном. Найденный вариант клонируется (т.е. размножается из единственного родоначального экземпляра).
За открытие этого механизма иммунолог из Японии Сусуму Тонегава получил в 1987 г. Нобелевскую премию (работа начата в Швейцарии, а завершена в США). Суть открытия в том, что ген может быть переделан в цитоплазме.
Указанный механизм рекомбинаций поставляет антитела, связывающие антигены довольно слабо. Для улучшения их «качества», для тонкой подстройки, осуществляется следующий этап, соматический (т.е. не связанный с размножением), – гипермутагенез. Включаясь после создания нужной комбинации фрагментов он осуществляет превращение «черновой болванки» в «готовое изделие».
Гипермутагенез заключается в том, что при клонировании гены «болванки» (первично найденного варианта) мутируют с огромной частотой (каждый тысячный нуклеотид заменяется, тогда как обычно точковый мутагенез в 100 миллионов раз менее интенсивен), а потом с их копий синтезируется масса чуть отличных друг от друга белковых цепей (рис. 1) антител, какое-то из которых оказывается подогнанным к антигену наилучшим образом. Этот окончательный вариант снова клонируется и запоминается клетками иммунной памяти, т.е. наследуется на время жизни особи (возникает приобретенный иммунитет).

Рис. 1. Избирательность гипермутагенеза (эффект Ву–Кэбота), которую установили в 1970 г. Тай Тэ Ву и Элвин Кэбот на легкой цепи одного из иммуноглобулинов мыши. (По Ройт и др., 2000.) Сходная картина получена и на других объектах. Вверху: 7 фрагментов белка с удачными заменами аминокислот. (Цифры справа – число необходимых для этого замен нуклеотидов.) Внизу: частота наблюдавшихся в опыте замен аминокислот
В этом, грубо говоря, состоит генетический принцип обеспечения разнообразия антител (термин Тонегавы): возникшие при перестройках фрагменты сшиваются (механизм Тонегавы), причем с нематричными вставками (механизм Альта–Балтимора, п.4), затем успешный вариант точно подгоняется к антигену (механизм гипермутагенеза), клонируется и запоминается (соматическое наследование).
Словом, гены антител образуются не за счет случайных мутаций, как думали прежде, а путем многостадийного процесса (рис. 2), в котором лишь одну ступень можно назвать мутагенезом и то в особом смысле: он направлен – в том смысле, что происходит только в нужных участках нужных генов, зато с неимоверной частотой.
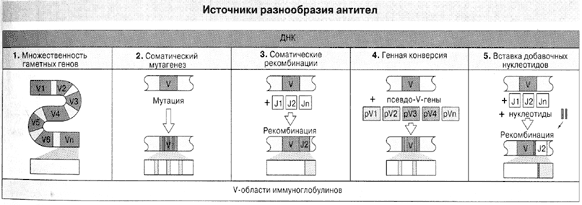
Рис. 2. Пять источников разнообразия антител ( По Ройт и др., 2000).
Кроме описанных в тексте, здесь указана генная конверсия, смысл которой тот же, что у рекомбинации по Тонегаве (комбинируются гипервариабельные участки V), но добавочными элементами комбинирования служат не соединительные сегменты J, а псевдогены, т.е. гены, утратившие собственную активность
Сам Тонегава в нобелевской лекции ясно провел параллель иммуногенеза с эволюцией: «Подобно организмам в экосистеме, эти лимфоциты – субъекты отбора антигенами, и приспособленнейший будет выживать. И . иммунную систему индивида можно рассматривать как своего рода Дарвинов микрокосм».
Параллель очевидна, но надо выяснить, причем тут Дарвин. Оказывается, иммуногенез схож не с естественным отбором, а с искусственным. Мы подошли к самому важному пункту.
6. Роль отбора в иммуногенезе
Мир учения Дарвина – это мир малых вариаций, последовательно вытесняющих друг друга в борьбе за дефицитные ресурсы. Только при таком понимании естественного отбора Дарвину удалось построить свою теорию. И хотя позже было предложено множество иных пониманий, годились они только для своих узких конкретных целей. Поэтому важно понять, насколько справедлива аналогия Тонегавы (не раз повторенная другими).