Типы рецепторов
У рыб есть механорецепторы боковой линии, расположенные вдоль туловища и на голове рыбы; они воспринимают движение воды относительно животного. Электрорецепторы — это другой тип рецепторов боковой линии. Во время эмбрионального развития все рецепторы боковой линии развиваются из того же участка нервной системы, что и слуховые и вестибулярные рецепторы. Так что слуховые рецепторы летучих мышей и электрорецепторы рыб — близкие родственники.
У разных рыб электрорецепторы имеют разную локализацию — они располагаются на голове, на плавниках, вдоль тела, а также и разное строение. Часто электрорецепторные клетки образуют специализированные органы. Мы рассмотрим тут один из таких органов, встречающихся у акул и у скатов,— ампулу Лоренцини. Лоренцини думал, что ампулы — это железы, вырабатывающие слизь рыбы. Ампула Лоренцини представляет собой подкожный канал, один конец которого открыт в наружную среду, а другой оканчивается глухим расширением; просвет канала заполнен желеобразной массой; электрорецепторные клетки выстилают в один ряд «дно» ампулы.
Интересно, что Паркер, который впервые заметил, что рыбы реагируют на слабые электрические токи, изучал и ампулы Лоренцини, но приписал им совсем другие функции. Он обнаружил, что, надавливая палочкой на наружный вход канала, можно вызвать реакцию акулы. Из таких опытов он сделал вывод, что ампула Лоренцини — это манометр для измерения глубины погружения рыбы, тем более, что по строению орган был похож на манометр. Но и на этот раз интерпретация Паркера оказалась ошибочной. Если акулу поместить в барокамеру и создать в ней повышенное давление,, то ампула Лоренцини на него не реагирует — и это можно нт>едвидетьх не ставя эксперимента: вода давит со всех сторон и никакого эффекта нет *). А при давлении только на пору в желе, которое ее заполняет, возникает разность потенциалов, подобно тому, как возникает разность потенциалов в пьезоэлектрическом кристалле.
Как же устроены ампулы Лоренцини? Оказалось, что все клетки эпителия, выстилающего канал, прочно соединены между собой особыми «плотными контактами», что обеспечивает высокое удельное сопротивление эпителия. Канал, покрытый такой хорошей изоляцией, проходит под кожей и может иметь длину в несколько десятков сантиметров. Напротив, желе, заполняющее канал ампулы Лоренцини, имеет очень низкое удельное сопротивление; это обеспечивается тем, что в просвет канала ионные насосы накачивают много ионов К+. Таким образом, канал электрического органа представляет собой отрезок хорошего кабеля с высоким сопротивлением изоляции и хорошо проводящей жилой.
«Дно» ампулы устилают в один слой несколько десятков тысяч электрореценторных клеток, которые тоже плотно склеены между собой. Получается, что рецепторная клетка одним концом смотрит внутрь канала, а на другом конце образует синапс, где выделяет возбуждающий медиатор, действующий на подходящее к ней окончание нервного волокна. К каждой ампуле подходят 10— 20 афферентных волокон и каждое дает много терминалей, идущих к рецепторам, так что в результате на каждое волокно действуют примерно 2 ООО рецепторных клеток.
Посмотрим теперь, что происходит с самими электро-рецепторными клетками под действием электрического поля.
Если любую клетку поместить в электрическом поле, то в одной части мембраны знак ГШ совпадет со знаком напряженности поля, а в другой окажется противоположным. Значит, на одной половине клетки МП возрастет, а на другой, наоборот, снизится. Получается, что всякая клетка «чувствует» электрические поля, т. е. является электрорецептором.
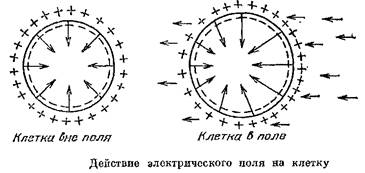
И понятно: ведь в этом случае отпадает проблема преобразования внешнего сигнала в естественный для клетки — электрический. Таким образом, электрорецепторные клетки работают очень просто: при надлежащем знаке внешнего поля деполяризуется синаптическая мембрана этих клеток и этот сдвиг потенциала управляет выделением медиатора.
Но тогда возникает вопрос: в чем особенности электрорецепторных клеток? Может ли выполнять их функции любой нейрон? Чему служит особое устройство ампул Лоренцини?
Да, качественно, любой нейрон может считаться электрорецептором, но если перейти к количественным оценкам, ситуация меняется. Естественные электрические поля очень слабы, и все ухищрения, которые использует природа в электрочувствительных органах, направлены на то, чтобы, во-первых, поймать на синаптической мембране возможно большую разность потенциалов, и, во-вторых, обеспечить высокую чувствительность механизма выделения медиатора к изменению МП.
Электрические органы акул и скатов обладают чрезвычайно высокой чувствительностью: рыбы реагируют на электрические поля напряженностью 0,1 мкВ/см. Так что проблема чувствительности решена в природе блестяще. Как же достигаются такие результаты?
Во-первых, обеспечению такой чувствительности способствует устройство ампулы Лоренцини. Если напряженность поля равна 0,1 мкВ/см, а длина канала ампулы равна 10 см, то на всю ампулу придется разность потенциалов в 1 мкВ. Практически все это напряжение будет падать на слое рецепторов, так как его сопротивление гораздо выше, чем сопротивление среды в канале. Акула тут прямо использует закон Ома: V = 11$, так как ток, текущий в цепи, один и тот же, то падение напряжения больше там, где выше сопротивление. Таким образом, чем длиннее канал ампулы и чем ниже его сопротивление, тем большая разность потенциалов подается на электрорецептор.
Во-вторых, закон Ома «применяют» и сами электрорецепторы; разные участки их мембраны тоже имеют разное сопротивление: синаптическая мембрана, где выделяется медиатор, имеет большое сопротивление, а противоположный участок мембраны — маленькое, так что и тут разность потенциалов распределяется возможно выгоднее,
Что же касается чувствительности синаптической мембраны к сдвигам МП, то она может объясняться разными причинами: высокой чувствительностью к сдвигу потенциала могут обладать Са-каналы этой мембраны либо сам механизм выброса медиатора. Очень интересный вариант объяснения высокой чувствительности выделения медиатора к сдвигам МП предложил А.Л. Вызов. Его идея состоит в том, что в таких синапсах ток, генерируемый постсинаптической мембраной, затекает в рецепторные клетки и способствует выделению медиатора; в результате возникает положительная обратная связь: выделение медиатора вызывает ПСП, при этом через синапс течет ток, а это усиливает выделение медиатора. В принципе, такой механизм обязательно должен действовать. Но и в этом случае вопрос является количественным: насколько эффективным является такой механизм, чтобы играть какую-то функциональную роль? В последнее время А.Л. Вызову и его сотрудникам удалось получить убедительные экспериментальные данные, подтверждающие, что такой механизм действительно работает в фоторецепторах.
Борьба с шумами
Итак, за счет разных ухищрений с использованием закона Ома на мембране электрорецепторов создается сдвиг потенциала порядка 1 мкВ. Казалось бы, что если чувствительность пресинаптической мембраны достаточно высока — а это, как мы видели, действительно так и есть,— то все в порядке. Но мы не учли, что повышение чувствительности всякого прибора вызывает новую проблему — проблему борьбы с шумами. Мы называли чувствительность электрорецептора, воспринимающего 1 мкВ, фантастической и теперь поясним, почему. Дело в том, что эта величина гораздо ниже уровня шумов.