Типы рецепторов
Итак, мы тут встретились еще с одним типом каналов — каналами, управляемыми изнутри клетки.
Если мы сравним фоторецептор позвоночного и беспозвоночного животного, то увидим, что в их работе очень много общего: имеется пигмент типа родопсина; сигнал от возбужденного пигмента передается к наружной мембране с помощью внутриклеточного медиатора; клетка не способна к генерации ПД. Различие же состоит в том, что внутриклеточный медиатор действует у разных организмов на разные ионные каналы: у позвоночных он вызывает гиперполяризацню рецептора, а у беспозвоночных, как правило,— деполяризацию. Например, у морского моллюска — гребешка — при освещении рецепторов дистальной сетчатки возникает их гиперполяризация, как у позвоночных, но механизм ее совершенно другой. У гребешка свет увеличивает проницаемость мембраны к ионам калия и МП сдвигается ближе к равновесному калиевому потенциалу.
Однако знак изменения потенциала фоторецептора не слишком существен, его всегда можно изменить в ходе дальнейшей обработки. Важно лишь, чтобы световой сигнал надежно преобразовывался в электрический.
Давайте рассмотрим для примера дальнейшую судьбу возникшего электрического сигнала в зрительной системе уже знакомых нам усоногих раков. У этих животных фоторецепторы при освещении деполяризуются и выделяют больше медиатора, но это не вызывает никакой реакции животного. Зато при затенении глаз рак принимает меры: убирает усики и т. д. Как же это происходит? Дело в том, что медиатор фоторецепторов усоногих раков тормозной, он гиперполяризует следующую клетку нейронной цепи, и она начинает выделять меньше медиатора, поэтому, когда свет становится ярче, никакой реакции не возникает. Наоборот, при затенении фоторецептора он выделяет меньше медиатора и перестает тормозить клетку второго порядка. Тогда эта клетка деполяризуется и возбуждает свою клетку-мишень, в которой возникают импульсы. Клетка 2 в этой цепи называется И-клеткой, от слова «инвертирующая», так как ее основная роль — менять знак сигнала фоторецептора. Усоногий рак имеет довольно примитивные глаза, да ему и немного надо; он ведет прикрепленный образ жизни и ему достаточно знать, что приближается враг. У других животных система нейронов второго и третьего порядков устроена гораздо сложнее,
В фоторецепторах рецепторный потенциал передается дальше электротонически и влияет на количество выделяющегося медиатора. У позвоночных или усоногих раков и следующая клетка безымпульсная и только третий нейрон цепочки способен к генерации импульсов. А вот в рецепторе растяжения наших мышц ситуация совершенно иная. Этот механорецептор представляет собой окончание нервного волокна, обвивающееся спиралью вокруг мышечного волокна. При растяжении ншпцы витки спирали, образованные безмиелиновой частью волокна, отходят друг от друга и в них возникает Г-цепторный потенциал — деполяризация, обусловленная открыванием Ка-каналов, чувствительных к деформации мембраны; этот потенциал создает ток, идущий через перехват Ранвье того же волокна, и перехват генерирует импульсы. Чем сильнее растянута мышца, тем больше рецепторный потенциал и тем выше частота импульсации.
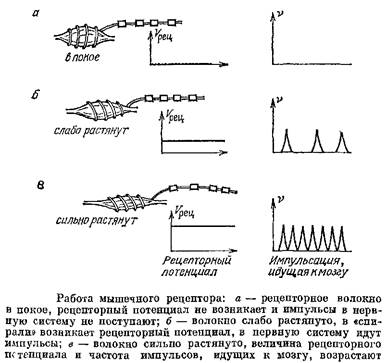
У этого механорецептора и преобразование внешнего воздействия в электрический сигнал, т. е. в рецепторный потенциал, и преобразование рецепторного потенциала в импульсы реализуется участком одного аксона.
Конечно, нам было бы интересно рассказать об устройстве разных рецепторов разных животных, ведь по своей конструкции и применению они бывают весьма экзотическими; однако каждый такой рассказ в конце концов сводился бы к одному и тому же: как внешний сигнал преобразуется в рецепторный потенциал, который управляет выделением медиатора или вызывает генерацию импульсов.
Но об одном типе рецепторов мы все же еще расскажем. Это электрорецептор. Его особенность состоит в том, что сигнал, на который надо реагировать, уже имеет электрическую природу. Что же делает этот рецептор? Преобразует электрический сигнал в электрический?
Электрорецепторы. Как акулы используют закон Ома и теорию вероятностей
В 1951г. английский ученый Лиссман изучал поведение рыбы гимнарха. Эта рыба обитает в мутной непрозрачной воде в озерах и болотах Африки и поэтому не всегда может для ориентации пользоваться зрением. Лиссман предположил, что эти рыбы, подобно летучим мышам, используют для ориентации эхолокацию.
Удивительная способность летучих мышей летать в полной темноте, не натыкаясь на препятствия, была обнаружена очень давно, в 1793г., т. е. почти одновременно с открытием Гальвани. Это сделал Лазаро Спалланцани — профессор университета в Павии. Однако экспериментальное доказательство того, что летучие мыши издают ультразвуки и ориентируются по их эху, было получено только в 1938 г. в Гарвардском университете в США, когда физики создали аппаратуру для регистрации ультразвука.
Проверив ультразвуковую гипотезу ориентации гимнарха экспериментально, Лиссман отверг ее. Оказалось, что гимнарх ориентируется как-то иначе. Изучая поведение гимнарха, Лиссман выяснил, что эта рыба обладает электрическим органом и в непрозрачной воде начинает генерировать разряды очень слабого тока. Такой ток не пригоден ни для защиты, ни для нападения. Тогда Лиссман предположил, что гимнарх должен обладать специальными органами для восприятия электрических полей — электросенсорной системой.
Это была очень смелая гипотеза. Ученые знали, что насекомые видят ультрафиолет, а многие животные слышат неслышимые для нас звуки. Но это было лишь некоторое расширение диапазона в восприятии сигналов, которые могут воспринимать и люди. Лиссман допустил существование совершенно нового типа рецепторов.
Ситуация осложнялась тем, что реакция рыб на слабые токи в это время была уже известной. Ее наблюдали еще в 1917 г. Паркер и Ван Хойзер на сомике. Однако эти авторы дали своим наблюдениям совсем другое объяснение. Они решили, что при пропускании тока через воду в ней меняется распределение ионов, и это влияет на вкус воды. Такая точка зрения казалась вполне правдоподобной: зачем придумывать какие-то новые органы, если результаты можно объяснить известными обычными органами вкуса. Правда, эти ученые никак не доказывали свою интерпретацию; они не поставили контрольного опыта. Если бы они перерезали нервы, идущие от органов вкуса, так чтобы вкусовые ощущения у рыбы исчезли, то обнаружили бы, что реакция на ток сохраняется. Ограничившись словесным объяснением своих наблюдений, они прошли мимо большого открытия.
Лиссман же, напротив, придумал и поставил множество разнообразных опытов и после десятилетней работы доказал свою гипотезу. Примерно 25 лет назад существование электрорецепторов было признано наукой. Электрорецепторы начали изучать, и вскоре они были обнаружены у многих морских и пресноводных рыб, а также у миног. Примерно 5 лет назад такие рецепторы были открыты у амфибий, а недавно — и у млекопитающих.
Где же расположены электрорецепторы и как они устроены?