Страница
2
В текальных пластинках есть специальные отверстия — поры для трихоцист. Трихоцисты располагаются по периферии клеток. Число их варьирует от немногих {Gymnodinium simplex) до сотен.
Трихоцисты состоят из узкой области — шейки, прикрепляющейся к теке в месте нахождения округлой поры с приподнимающимся краем, и более широкой части — тела; и шейка, и тело окружены одной мембраной. На поперечном срезе тело имеет квадратные или ромбические очертания и состоит из протеиновых субъединиц, упакованных в форме кубической решетки, шейка же образована пучком спирально закрученных фибрилл. Выбрасываемые при возбуждении через поры в теке трихоцисты значительно длиннее и уже. Они представляют собой белковые нити с периодической исчерченностью под прямым углом к их длинной оси. Есть данные, что примордиальные трихоцисты возникают в центральной части клетки в везикулах — дериватах аппарата Гольджи.
Жгутиковый аппарат. Два жгутика различаются по форме: один «продольный», обычного строения, содержит ак-сонему 9 — 9 + 2, на мембране его во многих случаях в два ряда располагаются волоски; второй — «поперечный» — заключает внутри растянувшейся мембраны, помимо аксонемы, еще исчерченный тяж; мембрана его несет односторонний ряд длинных тонких волосков. Как и у других водорослей, мастигонемы возникают интрацеллюлярно: везикулы, содержащие промастигонемы, в цитоплазме можно видеть часто вблизи ядра и в тесной связи с дикгиосомами и эндоплазма-тическим ретикулумом.
Пузулы. Большинство монадных динофитовых водорослей обладает вместо отсутствующих пульсирующих вакуолей особой органеллой — пузулой. Пузула представляет собой инвагинацию цитоплазматической мембраны в форме мешка или трубки, глубоко вдающуюся внутрь клетки и непосредственно соприкасающуюся с вакуолью клеточного сока. Вакуоль окружает пузулу, стенка которой становится таким образом двух-мембранной. Пузулы изливают свое содержимое наружу через жгутиковые каналы. В случае крупной мешковидной пузулы поверхность контакта между плазмалеммой и тонопластом увеличивается благодаря инвагинациям разветвлений вакуоли внутрь пузулы. То же относится и к трубковидным пузулам. Если приходящее в контакт с вакуолярной системой впячивание плазмалеммы сообща-

ется с наружной средой не непосредственно, а с помощью собирающего пространства, говорят о собирательной пузуле. Расположение в клетке пузырей, собирательных камер, выводковых трубок у разных представителей варьирует. Часто около мешковидной находится собирательная пузула, изливающаяся или в мешковидную, или через свой собственный проток.
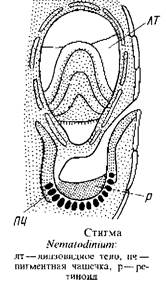
Глазки, расположенные вне хлоропластов у основания жгутиков, также отличаются большим разнообразием: от простейшего глазка в виде массы глобул {Woloszynskia coronata) до сложной светочувствительной органеллы, состоящей из стекловидной, преломляющей свет линзы и пигментного тела, например у рода эритропсис. У нематодиниума стигма тоже состоит из прозрачного линзовидного тела, окружающей его снизу пигментной чашечки и расположенного на дне этой чашечки светочувствительного ретиноидного тела. Линза точно фокусирует свет на дне пигментной чашечки. С физической точки зрения была доказана возможность получения изображения объекта на ретиноидиом слое.
Размножение вегетативное, бесполое, половое. [',,
Отдел Dinophyta делится гта три класса: десмофициевые, динофициевые и бласто-динофициевые. Представители класса дес-мофициевых — монадные, голые или одетые панцирем, преимущественно морские формы со жгутиками, отходящими от переднего конца клетки. Класс динофициевые объединяет монадные, ризоподиальные, паль-меллоидные, коккоидные и нитчатые формы. Монадные клетки дорзивентраль-ные, с поперечной и продольной ложбинками, в которых располагаются жгутики, отходящие в месте пересечения ложбинок на вентральной стороне клетки. Класс бластодинофициевые — искусственная группа, объединяющая паразитные формы.
КЛАСС ДЕСМОФИЦИЕВЫЕ DESMOPHYCEAE -
Представителем может служить преимущественно морской род пророцентрум, иногда размножающийся столь обильно, что вызывает цветение прибрежных вод. Клетки более или менее овальные, с боков несколько уплощенные, лишены ложбинок или бороздок. Две главные пластинки тек блюдцевидные, смыкаются вдоль всего края, за исключением их переднего конца, где в одной из них имеется выемка. В апикальной обласги располагается ряд мелких толстых пластинок, довольно прочно соединенных друг с другом так, что апикальная структура имеет тенденцию отделяться как одно целое. От апикальных пластинок отходит и характерный для рода апикальный шип, или зубец,— трубчатая или сдвоенная структура. Главные текальные пластинки гладкие или орнаментированные, перфорированы рядом трихоцистных пор. На переднем конце клетки имеются два жгутика, выходящие через индивидуальные каналы в апикальных пластинках. Один жгут направлен вперед, превышает по длине клетку, имеет заостренный конец и, по-видимому, не несет волосков. Второй жгут, несмотря на то что клетка лишена каких-либо следов поперечной борозды, свойственной монадным клеткам динофициевых водорослей, является по структуре типичным «поперечным» жгутом. Он закручен и внутри свободной мембраны, помимо аксонемы, содержит исчерченный тяж; мембрана его несет односторонний ряд мелких волосков. Основания жгутов в отличие от динофлагеллят параллельны друг другу и располагаются каждое в цитоплазматическом выступе в полости под текой. При вхождении жгутика в цитоплазматический выступ находится толстый трехслойный базальный диск, замыкающий центр аксонемы, под которым периферические дублеты превращаются в триплеты. Оба жгутика различаются по расположению и структуре своих корней. Один жгутик в месте выхода из цитоплазмы имеет «воротничок», и несколько микротрубочек тянутся от основания жгутика к «воротничку». У другого жгута проксимальный конец базального тела заключен в электронно-плотный материал, от которого отходят вперед трубчатый корень и корень с поперечной исчерченностью.
В протопласте находится один желтовато-бурого цвета париетальный сетчатый хлоропласт, окруженный трехмембранной оболочкой и содержащий ламеллы из двух-трех тилакоидов. Пиреноид ограничен растяжением оболочки хлоропласта, матрикс его обычно пересечен несколькими двухтилакоидными ламеллами. Гранулы крахмала откладываются вне хлоропласта. В задней половине клетки расположено ядро, более или менее сферическое, с ядрышком, оболочка ядра с многочисленными порами. Деление ядра такое же, как у других динофлагелля г: ядро становится перфорированным туннелями, которые содержат многочисленные микротрубочки и ориентированы большей частью поперек ядра — под прямым углом к плоскости деления. Тела Гольджи, митохондрии,

мешковидная пузула — как и у других динофитовых водорослей. Предшественники трихоцист находятся вблизи центра клетки, а зрелые трихоцисты — под текой. Размножение осуществляется путем продольного деления клетки, причем дочерние протопласты получают одну половину материнского панциря, а другую выделяют заново.