Отдел диатомовые водоросли

цитоплазмы может перейти во внутреннюю щель этой же ветви шва, чтобы повторить пройденный путь, но может попасть и непосредственно в полость клетки или же через горизонтальную борозду, соединяющую в центральном узелке обе ветви шва, переходит во внутреннюю щель другой ветви шва и начинает там круговой путь в противоположном направлении. Трение струящейся цитоплазмы о субстрат и развивает моторную силу, движущую клетку в направлении, противоположном току цитоплазмы. Движение форм, имеющих каналовидный шов без центрального узелка, значительно интенсивнее благодаря непрерывному току цитоплазмы.
Согласно другой гипотезе скольжение диатомовых обусловлено движением жидкости в шве, представляющем собой пластинчатый капилляр, в который жидкость втягивается под действием капиллярных сил.
У имеющих шов пеннатофициевых хлоропласты располагаются на поисковой стороне клетки, лишь слегка заходя на створковую. Перед делением клетки хлоропласты перемещаются на створку. У бесшовных форм хлоропласты располагаются со стороны створки.
Ничего сравнимого со швом не наблюдается у представителей центрофициевых. Они все неподвижны.
Многие диатомей в течение всей жизни встречаются в виде отдельных клеток, однако известны и колониальные формы, как свободноплавающие, так и прикрепленные. Колонии берут свое начало от одной клетки, которая в результате ряда делений дает свойственную данному виду форму колонии. Как правило, колонии образуются у видов, не способных к движению, т. е. совсем не имеющих шва. Виды же с вполне развитым щелевидным или каналовид-ным швом формируют колонии редко, и при этом клетки, входящие в состав колонии, обычно не теряют способности к движению.
Клетки прикрепляются к субстрату и соединяются в колонии с помощью слизи, выделяемой через сравнительно крупные поры на поверхности створки. Иногда в скреплении клеток в колонии, помимо слизи, принимают участие различные выросты створок—рога, щетинки, зубчики и пр. Все громадное разнообразие форм колоний можно свести к трем основным типам. К первому типу относятся студенистые колонии, в которых клетки целиком погружены в выделяемую ими слизь. Колонии имеют вид бесформенных комочков или трубок, неэетвящихся или разнообразно разветвленных.
Второй тип включает нитевидные, лентовидные, цепочковидные и звездчатые колонии, форма их зависит от способа скрепления клеток в колонии и от формы створок. Во всех случаях клетки соединяются только створками, соединения поясками никогда не бывает, так как в плоскости пояска клетки делятся. Если смежные клетки соединяются всей поверхностью створки, то в зависимости от формы створок образуются колонии нитевидные, как у некоторых видов циклотеллы и у рода мелозира, лентовидные— например у фрагилярии, вееровидные — у меридион. Если же клетки смыкаются только концами створки, то возникают или зигзагообразные цепочки, как у рода диатома, или звездчатые колонии, характерные для рода астерионелла
У различных планктонных центрофициевых клетки соединяются выростами створок, слизистыми тяжами и тогда образуются не плотные, а рыхлые цепочки, в которых клетки более или менее удалены друг от друга. Таковы роды хетоцерос, скелетонема {Skeletonema, рис. 27, А—В), талассиози-ра.
Третий тип колоний наблюдается у форм, прикрепленных к субстрату слизью, которая выделяется локально на базальном конце клетки через поры. Слизь имеет вид тяжей неразветвленных или древовидно ветвящихся, на концах которых сидят клетки — по одной или помногу. Примерами могут служить виды синедры, гом-фонемы, ликмофоры.
Электронно-микроскопические исследования панциря диатомовых водорослей показали, что кремневые стенки панциря бывают однослойными и двухслойными независимо от принадлежности данного вида к центрофициевым или пеннатофициевым диатомовым. В первом случае стенка панциря состоит из одной пластины, во втором—из двух пластин: наружной и внутренней, пространство между которыми разделено вертикальными перегородками на камеры. Кроме того, было показано, что створки панциря всегда пористые, и один из основных элементов структуры панциря — именно поры. По классификации X. Окуно различают три основных типа пор: отверстия, локулы, альвеолы. Отверстия — это мелкие, в виде следа булавочного укола округлые или щелевидные перфорации, возникающие в более или менее тонких однослойных стенках. Отверстия лишены замыкающих ситовидных мембран. В двухслойных стенках встречаются или округлые и полигональные перфорации локулы, или более или менее вытянутые перфорации— альвеолы. И те и другие снабжены замыкающими ситовид-
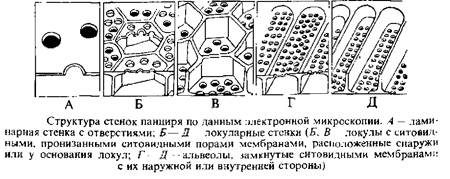
иыми мембранами, перфорированными, в свою очередь, ситовидными порами. Локулы могут иметь хорошо выраженные высокие латеральные стенки, которые вдаются глубоко внутрь, и тогда ситовидная мембрана находится снаружи локулы; если латеральные стенки высоко выступают наружу, то ситовидная мембрана расположена у основания локулы. В первом случае изнутри, а во втором--снаружи локул обычно наблюдается покровная мембрана, перфорированная всегда одной крупной порой— покровной порой. В том и другом типе локул покровная мембрана может отсутствовать. Латеральные стенки бывают с латеральными порами или без них. В толстых стенках, перфорированных удлиненными альвеолами, как и в случае локул, латеральные стенки вдаются внутрь или выступают кнаружи, соответственно ситовидная мембрана располагается снаружи от альвеолы или у ее основания. Покровные мембраны могут присутствовать или отсутствовать. Пор в латеральных стенках альвеол до сих пор не наблюдалось.
Ситовидные мембраны, которые замыкают локулы или альвеолы на их наружной или внутренней поверхностях, имеют характерную тонкую структуру. Ситовидная мембрана перфорирована одной или многими крошечными ситовидными порами округлых, эллиптических, прямоугольных или полигональных очертаний. Ситовидные поры, в свою очередь, часто замкнуты тонкой вторичной мембраной с ультратонкими вторичными ситовидными порами. Форма, размер и расположение ситовидных пор могут служить в качестве надежного признака для идентификации видов.
Несколько подробнее можно рассмотреть структуру панциря у рода пиннулярия. В панцире пиннулярии альвеолы поперечно вытянуты, простираясь почти от шва до края створки. Они представлены корытообразной полостью, замкнутой снаружи только пористой ситовидной мембраной. У некоторых видов пиннулярии альвеолы, лишенные покровной мембраны, полностью открыты внутрь, у других видов альвеолы изнутри загорожены тонкой покровной мембраной с отверстием— покровной порой. Покровные поры могут быть округлыми, эллиптическими или другой формы, они расположены на створке в две продольные линии. Эти ряды покровных пор различимы и в световом микроскопе. Поперечные боковые стенки альвеол более или менее глубоко вдаются внутрь и лишены пор. Таким образом, соседние альвеолы полностью разделены этими боковыми стенками.
Электронно-микроскопическая тонкая структура пор панцирей диатомовых водорослей строго видоспецифична. Согласно X. Оку-но, «надежность тонкой структуры пор панциря для идентификации видов диатомовых водорослей может быть сравнима с надежностью отпечатков пальцев для идентификации человеческих индивидуумов».