Механизмы дыхания растений
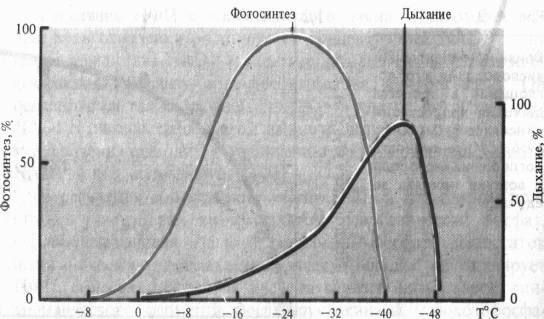
Для дыхания каждого вида растений и его органов существуют определенные минимальные (когда дыхание еще возможно), оптимальные и максимальные температуры. В отличие от фотосинтеза дыхание у зимующих растений наблюдается при очень низких температурах (у хвои ели и сосны при — 25 °С). Температурный оптимум его у большинства видов умеренных широт лежит в пределах 35 — 40 °С, т. е. на 5 — 10 °С выше, чем для фотосинтеза. Максимальные температуры (45 — 55 °С) определяются способностью белков к денатурации.
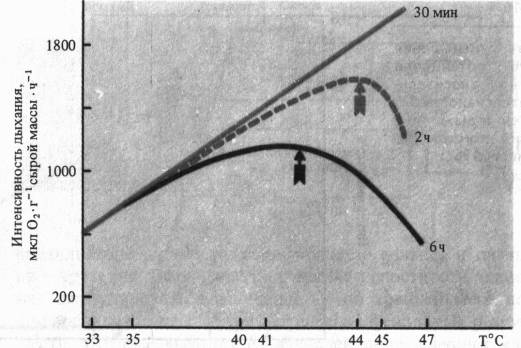
При рассмотрении влияния на дыхание крайних температур важно учитывать продолжительность действия этого неблагоприятного фактора. Так, при кратковременных экспозициях (30 мин) температурный оптимум дыхания листьев гречихи сахалинской может быть очень высок (> 47 °С), тогда как при двухчасовом действии оптимум смещается к 44 °С, а при шестичасовом—до 42 °С.
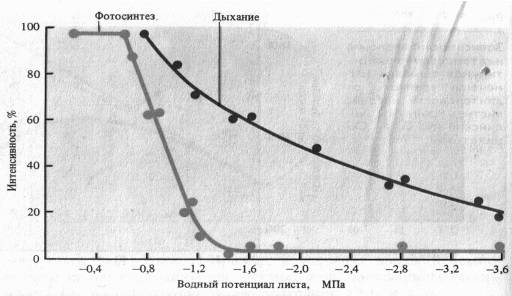
Водный режим. Изменение оводненности тканей растения отражается на интенсивности дыхания. В листьях проростков при быстрой потере воды вначале отмечается усиление дыхания — обычная кратковременная реакция на раздражение. При постепенном снижений оводненности этого не происходит. Продолжающийся водный дефицит вызывает постепенное снижение дыхания, причем более медленное, чем падение интенсивности фотосинтеза. Более раннее прекращение фотосинтетической активности при водном дефиците является следствием закрытия устьиц (уменьшение поступления С02 в лист) и потери тургорного давления в клетках.
Необходимость определенной степени оводненности тканей для процесса дыхания отчетливо проявляется при изучении дыхания семян. Дыхание воздушно-сухих семян (10—11% Н20) ничтожно мало. При повышении влажности семян до 14—15% дыхание возрастает в 4 —5 раз, а при дальнейшем повышении оводненности семян до 30 — 35% дыхание увеличивается в тысячи раз. При этом важную роль играет температура, при которой хранятся семена: при 0— 10°С влияние влажности на интенсивность дыхания значительно меньше, чем при 18 — 25 °С. Резкий подъем дыхания набухших семян сопровождается значительным выделением тепла, что может привести к их перегреву («самовозгоранию») при хранении.
Реакция растительных тканей на потерю воды зависит также от возраста, физиологического состояния, экологической принадлежности организма.
Минеральные вещества. Добавление раствора солей в воду, на которой выращивают проростки, обычно усиливает дыхание корней. Этот эффект получил название «солевого дыхания». Полагают, что солевое дыхание отражает усиление обмена веществ, необходимое для энергетической поддержки активного транспорта ионов корнем. Однако до конца это явление не изучено. Отмечены случаи, когда увеличенная интенсивность дыхания сохранялась после удаления солей. В тканях других органов этот эффект удается получить не всегда.
Свет. Действие света на дыхание зеленых органов растений из-за методических трудностей изучено недостаточно, так как одновременно с дыханием в них осуществляется фотосинтез. Освещенность, при которой интенсивность фотосинтеза равна интенсивности дыхания (по уровню поглощенного и выделенного С От), называется компенсационным пунктом.
При работе с незелеными растениями или с незелеными органами растений возможно непосредственное изучение действия света на дыхание. Дыхание (поглощение 02 и выделение С02) незеленых тканей активируется светом коротковолновой части спектра — ближним ультрафиолетом (380 нм), синими и зелеными лучами (400 — 500 нм) .
В области 380 — 600 нм лежат максимумы поглощения каротиноидов, флавинов и всех цитохромов дыхательной цепи митохондрий. Усиление дыхания может быть связано с прямым действием света на эти компоненты окислительно-восстановительных систем клеток.
Остается нерешенным вопрос о характере взаимодействия процессов фотосинтеза и дыхания в зеленых клетках в ходе фотосинтеза. Обсуждаются две точки зрения об использовании АТР, образующегося при фотофосфорилировании. Согласно одной из них на свету хлоропласта большую часть синтезируемой при фотофосфорилировании АТР используют на синтез углеводов, отдавая на нужды клетки лишь небольшую ее часть. АТР в цитоплазму поставляют митохондрии, которые используют ассимиляты. В темноте хлоропласта сами нуждаются в поступлении энергии из цитоплазмы.
По другой гипотезе АТР фотосинтеза на свету обеспечивает энергией не только фиксацию С02, но и другие энергозависимые процессы в клетке. Окислительное фосфорилирование в митохондриях на свету резко замедляется, а цикл Кребса ускоряется, так как его интермедиаты необходимы для функционирования хлоропластов.
Список использованной литературы
1.В.В. Полевой Физиология растений М.: Высшая школа 1989г.
2.Н.И. Якушкина Физиология растений М.: Просвещение 1993г.