Закономерности изменчивости и наследственности
Так, у душистого горошка есть ген А, обуславливающий синтез бесцветного предшественника пигмента – пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотиром аа ВВ и Аа вв имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором – есть пропигмент, но нет фермента, переводящего пропигмент в пигмент. При скрещивании двух растений душистого горошка с белыми цветками (Аавв х ааВВ) получается гибрид с пурпурной окраской цветков (АаВв)
Эпистатическое взаимодействие неаллельных генов противоположно комплиментарному действию генов. Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой. Гены, подавляющее действие других неаллельных генов, называются супрессорами, или подавителями. Они могут быть как доминантными, так и рецессивными. Так,окрашенными плоды тыквы будут только в том случае, если в генотипе растений отсутствует доминантный ген из другой аллельной пары. Этот ген подавляет развитие окраски плодов тыквы, а его рецессивная аллель не мешает окраске развиватся.
Полимерия. В некоторых случаях установлено, что проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие. Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зёрна, растения а1а1а2а2 – белые зёрна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску. Таким образом, накопление определённых аллелей в генотипе может вести к изменению выраженности признаков.
Генетика пола.
Пол – это совокупность морфологических, физиологических, биохимических и других признаков организма, обусловливающих воспроизведение себе подобного. При изучении наборов хромосом мужских и женских особей обратили внимание на тот факт, что у большинства женских организмов все хромосомы образуют пары, а у мужских помимо парных (гомологических) хромосом имеются две непарные. В дальнейшем было установлено, что эти непарные хромосомы как раз и определяют пол организма. Большая из непарных хромосом, которая содержится в женском кариотипе в двойном наборе, а в мужском – в одиночном, названа X– хромосомой. Меньшая из непарных хромосом, которая содержится только у особей мужского пола, названа Y– хромосомой. Парные хромосомы, одинаковые у мужского и женского организма, называются аутосомами, а X - и Y- хромосомы – половыми (гетерохромосомами).
Благодаря скрещиванию и комбинации генов возникают генотипы, позволяющие видам организмов успешно приспосабливаться к меняющимся условиям внешней среды. Половой процесс обеспечивает широкое распространение внутри вида любого гена, особенно
имеющего приспособительное значение.
Известно несколько способов определения пола у животных. Важнейший из них - определение пола в момент оплодотворения. В этом случае большую роль играет хромосомный
аппарат зиготы.
В диплоидном наборе у человека содержится 46 хромосом (23 пары): 22 пары аутосом и одна пара половых хромосом.
У женского организма все хромосомы парные, это две X – хромосомы, а у мужского – X и Y. Набор хромосом женщины может быть представлен следующей записью: 44А+2X, а мужчины – 44А+XY.
Пол, имеющий две одинаковые половые хромосомы (XX), называется гомогаметным, так как он образует только один тип гамет, содержащих X – хромосому. Пол, определяемый различными половыми хромосомами (XY), называется гетерогаметным, так как образует два типа гамет, содержащих X и Y – хромосомы соответственно. Пол будущего организма определяется в момент оплодотворения и зависит от того, какой из сперматозоидов оплодотворит яйцеклетку. При оплодотворении яйцеклетки сперматозоидом, содержащим X – хромосому, в зиготе будет две X – хромосомы и из неё разовьётся женский организм. При оплодотворении яйцеклетки сперматозоидом с Y – хромосомой в зиготе будет содержаться X – и Y – половые хромосомы, и она даст начало мужскому организму. Нетрудно заметить, что образование сперматозоидов с X – Y – хромосомами равновероятно и, следовательно, механизм гаметогенеза определяет, не только пол, но и примерное численное равенство полов в каждом поколении.
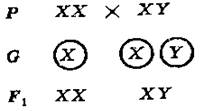
У всех млекопитающих, человека и мухи дрозофилы гомогаметным является женский пол, а гетерогаметным – мужской. У птиц и бабочек, наоборот, гомогаметен мужской пол, а женский – гетерогаметен. У некоторых насекомых женский пол содержит две X – хромосомы (XX), а мужской – одну (XO), т. е. в кариотипе самцов отсутствует Y – хромосома.
В половых хромосомах помимо генов, определяющих пол организма, содержатся и другие, не имеющие отношения к полу. Например, только в X – хромосоме содержатся гены, определяющие нормальную свёртываемость крови (доминантный признак, Н) и несвёртываемость – гемофилию (рецессивный признак, h), нормальное цветоощущение (доминантный признак,С ) и цветовую слепоту – дальтонизм (рецессивный признак, с). Так как аномальные признаки рецессивны и поэтому у женщин не проявляются. В случаях, когда эти гены несёт одна из Х – хромосом, такая женщина оказывается и передаёт их с Х – хромосомой своим сыновьям, у которых аномалии проявятся, так как Х- хромосома у них одна.
Ген, обусловливающий нормальную свёртываемость крови, - Н, а ген, при котором кровь теряет способность свёртывания, - h.
Женщина передаёт половине своих сыновей Х – хромосому с геном нормальной свёртываемости крови, а половине – Х – хромосому с геном гемофилии. Среди её сыновей могут быть и здоровые и гемофилики.
Признаки, определяемые генами, локализованными в негомологичном участке Х – хромосомы, называются сцепленными с полом, а локализацию генов в половой хромосоме –
сцеплением генов с полом. Распределение этих генов в потомстве должно соответствовать распределению половых хромосом в мейозе и их сочетанию при оплодотворении. При записи браков по признакам, сцепленным с полом, соответствующий ген обозначают в виде индекса:
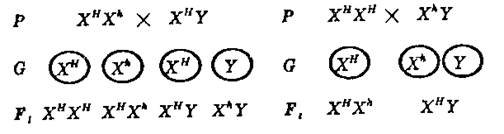
Сцепленные с полом признаки передаются от матери к дочерям и сыновьям, а от отца – только к дочерям. Следует подчеркнуть, что у мужских организмов единственный рецессивный ген, содержащийся в X – хромосоме, всегда проявляется (XhY), так как в Y – хромосоме нет аллельного гена.
Основные положения хромосомной теории наследственности.
- Гены расположены в хромосомах линейно в определённых локусах (участках). Аллельныегены гены занимают отдельные локусы в гомологичных хромосомах.
- Гены одной хромосомы образуют группу сцепления; число их равно гаплоидному набору хромосом.
- Между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер).
- Расстояние между генами пропорционально проценту кроссинговера между ними и выражается в морганидах (1морганида равна 1%кроссинговера).