Страница
3
Где происходит рекомбинация
Мы уже говорили, что рекомбинация - это случайный процесс перетасовки генов. Однако сами точки рекомбинации распределены по геному далеко не случайно (рис.7), а согласно ряду строгих правил.
Правило обязательного обмена гласит: всякая пара хромосом да имеет хотя бы один обмен. Общее число обменов на хромосому зависит от ее размера. На больших хромосомах бурозубки (af и bс) их может быть пять или шесть, но даже самые маленькие хромосомы (например, tu), как правило, имеют хотя бы один обмен.
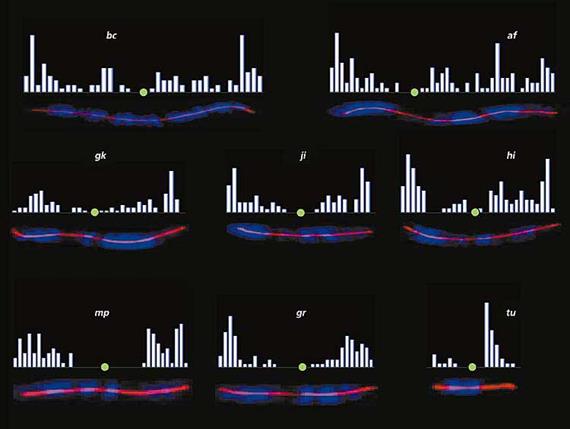
Рис.7. Распределение точек рекомбинации по хромосомам обыкновенной бурозубки.
Это правило обусловлено тем, что рекомбинация в мейозе, кроме генетического смысла (перетасовки генов), несет и чисто механическую функцию: образование и сохранение физической связи между гомологами вплоть до их расхождения в первом делении мейоза. Если между парой гомологов не произошло ни одного обмена, то они расходятся не как положено (один к одному полюсу деления, другой - к другому), а как попало, т.е. оба гомолога могут уйти в одному полюсу. Тогда у одних гамет будет избыточная доза генов, а у других этих генов не будет вовсе. И то, и другое чаще всего ведет к гибели организмов.
Правило теломерного пика: обмены могут быть в любых районах хромосом, но чаще всего они локализуются на самом краю хромосом - вблизи теломер. Это обусловлено, видимо, механикой рекомбинации, а не ее генетическим смыслом. Я уже говорил, что сближение хромосом начинается с того, что их концы собираются в одной точке на ядерной мембране, т.е. самый первый контакт между гомологами происходит именно на краях хромосом, и эти самые края находятся в тесном контакте дольше, чем все остальные районы. Там же возникают первые опознавательные связки на уровне ДНК, и времени на образование и разрешение этих связок отводится больше, чем в других точках хромосом.
Я с удовольствием вспоминаю эксперимент, который мы с И.П.Горловым провели еще в начале 1990-х годов [7]. Нас интересовал вопрос: что определяет частоту рекомбинации в каждом районе хромосомы - его положение или генетическое содержание? Мы сравнили распределение обменов на первой хромосоме у нормальных мышей и у гомозигот по инверсии терминальной половины этой хромосомы. Если важно положение, то распределение должно быть одинаковым; если важно содержание, то у гомозигот по инверсии и распределение должно быть инвертированным. Полученные нами распределения были одинаковыми (рис.8). Следовательно, решающую роль играет положение.

Рис.8. Распределение точек рекомбинации в хромосоме 1: слева - у нормальных мышей; в середине - у мышей, гомозиготных по инверсии в этой хромосоме, справа - распределение, которое было бы, если бы оно контролировалось не позицией, а генетическим содержанием района хромосомы.
Это механическое правило может иметь очень глубокий генетический и эволюционный смысл. Инверсии - одни из самых частых хромосомных перестроек, которые встречаются в природных популяциях и/или отличают виды друг от друга. Можно думать, что естественный отбор способствует фиксации инверсий, переносящих на края хромосом (т.е. в рекомбинационно горячие зоны) именно те гены, которые нужно часто перетасовывать, и наоборот, забрасывают в холодные зоны те блоки генов, которые лучше не трогать.
Правило интерференции: два обмена редко возникают в непосредственной близости друг от друга. Создается впечатление, что уже возникший обмен мешает (interfere) другому обмену возникнуть рядом. Это отталкивание обменов друг от друга получило название интерференции. Существует очень большой разброс в дистанциях между соседними обменами: от 1 мкм (т.е. на пределе разрешающей способности микроскопа) до 10 и более, а средние дистанции между соседними обменами у бурозубки составляют 5-6 мкм. Именно на этом расстоянии от теломерного пика обменов возникает второй пик. Если хромосома достаточно длинная, то примерно на таком же расстоянии возникнет третий пик, и так далее. Замечательно, что если мы обнаруживаем на большой хромосоме один-единственный обмен, то он, как правило, находится достаточно далеко от края. Почему? Потому что интерференция этого обмена не оставляет места для других обменов.
Парадоксально, но лучше всего явление интерференции интерпретируется в грубо механических терминах. Если представить хромосомы в виде упругих шлангов, скрученных друг с другом, становится понятно, что два перегиба рядом не возникают. Но в наше молекулярное время такие сантехнические аналогии выглядят по меньшей мере неприлично. Должны быть какие-то молекулярные механизмы интерференции обменов. По молекулярным понятиям несколько микрометров - это гигантское расстояние.
Существует несколько гипотез о механизмах интерференции, но ни одна из них не является общепринятой и ни одна из них мне не нравится. Поэтому я лучше приведу предложенную И.П.Горловым [8] гипотезу о том, зачем нужна интерференция. Суть ее такова. Ничего в природе не бывает бесплатно. Конечно, рекомбинация дает определенные преимущества, но она обходится и организму недешево. Чего стоит одно разрезание живой ДНК! Естественный отбор должен балансировать рекомбинацию таким образом, чтобы ее преимущества превышали затраты. Два обмена стоят дороже, чем один, где бы они ни были расположены. Но если мы подумаем о преимуществах, то станет понятно, что чем ближе они друг к другу, тем менее полезны в смысле перетасовки генов. Второй обмен в непосредственной близости от первого практически аннулирует его эффект, возвращая гомологичные хромосомы в исходное состояние (рис.9). Поэтому, если уж тратиться на обмены, то надо расположить их подальше друг от друга. Что и происходит благодаря интерференции, только до сих пор непонятно как.
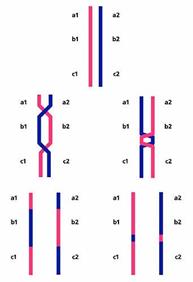
Рис.9. Результаты двух рекомбинационных событий на далеком (слева) и близком (справа) расстоянии друг от друга. В первом случае рекомбинантные хромосомы отличаются от родительских по набору генов a, b, c; во втором - не отличаются.
Все перечисленные выше правила так или иначе обусловлены позиционными эффектами, которые играют, видимо, главную роль в распределении обменов по хромосомам.
Правило светлого района связано с особенностями упаковки ДНК мейотических хромосом. Отдельные районы хромосом сильно отличаются по этому признаку. Хромосомы млекопитающих после обработки красителями, специфично связывающимися с ДНК, выглядят поперечно-полосатыми. В интенсивно окрашенных (темных) участках концентрация ДНК выше, чем в светлых. Особенности упаковки зависят от генетического состава ДНК. Сильно огрубляя ситуацию, можно сказать, что гены сосредоточены в основном в светлых районах, а темные главным образом состоят из бессмысленных и сильно повторенных последовательностей ДНК. Но это очень грубая картина. Гены есть и в темных районах, а в светлых довольно много бессмысленных последовательностей.