Страница
2
Мощное и радикальное средство борьбы с храповиком Мёллера - рекомбинация. Перетасовывая гены при образовании гамет, она может перегрузить мутациями одни гаметы и одновременно недогрузить другие. В итоге особи, возникшие из перегруженных мутациями гамет, погибают, а продукты гамет, очищенных от мутаций, процветают (рис.2,б). Это позволяет рекомбинирующим организмам избавиться от ограничений, накладываемых храповиком Мёллера. Они могут позволить себе роскошь иметь большие геномы. Отсюда получается, что все мы высшие и сложные оттого, что наши далекие одноклеточные предки открыли для себя рекомбинацию и создали механизмы, гарантирующие регулярную перетасовку генов из поколения в поколение.
Гипотеза Мёллера - не единственное объяснение преимуществ рекомбинации. Очень подробные обзоры гипотез о преимуществах рекомбинации даны в книгах Дж.Мэнард Смита [1] и М.Ридли [2].
Как происходит рекомбинация
У всех эукариот рекомбинация происходит в мейозе [3]. Этот процесс изучают в школе, и по степени вызываемой у учащихся ненависти мейоз может соперничать разве что с двойным оплодотворением у растений по Навашину. Что обидно, поскольку процесс этот по-своему красив (рис.3).
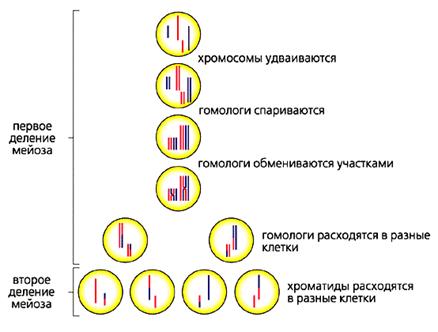
Рис.3. Схема мейоза.
Красным обозначены материнские хромосомы, синим - отцовские.
Некоторые сравнивают мейоз с танго [4]. В первом туре мейоза пары удвоенных гомологичных хромосом (одна получена от отца, другая от матери) находят друг друга, тесно свиваются, обмениваются участками, а потом расходятся в разные стороны, в разные клетки, чтобы уже не встретиться никогда. Второй тур мейоза менее романтичен. В нем гомологи выстраиваются вдоль экватора клетки и их хроматиды расходятся к разным полюсам. Так из одной генеративной клетки получаются четыре гаметы с разными наборами генов.
Рекомбинация происходит в первом, романтическом туре мейоза. Рассмотрим его более подробно. Сближение гомологичных хромосом начинается с того, что их концы, скользя по ядерной мембране, собираются в одной точке и формируется структура, романтически названная букетом. В нем гомологичные хромосомы оказываются поблизости друг от друга и приступают к взаимному опознанию. Оно, видимо, идет в два этапа: сначала приблизительное, а затем точное. Приблизительное опознание может происходить по принципу штрих-кодов. Известно, что хромосомы представляют собой комплекс ДНК с белками, набор которых и характер связывания с ДНК во многом определяются последовательностью нуклеотидов. Поэтому каждая хромосома отличается индивидуальным, только для нее специфичным распределением белков - штрих-кодом. Поскольку гомологичные хромосомы в основном сходны по последовательностям ДНК, они должны иметь сходные штрих-коды. Таким образом, грубое распознавание может быть достигнуто простым их совмещением.
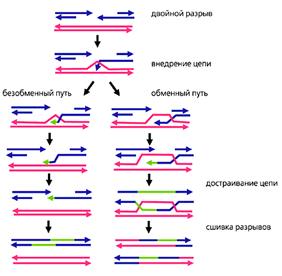
Рис.4. Молекулярные механизмы тонкого опознавания и рекомбинации гомологичных хромосом.
Красным обозначена ДНК материнской хромосомы, синим - отцовской, зеленым - ДНК, достроенная в ходе репарации разрывов.
Обратите внимание, что достройка идет по матрице ДНК гомолога, а не по собственной матрице [5].
Процесс тонкого опознания начинается с того, что в ДНК мейотических хромосом возникают множественные двунитевые разрывы (рис.4). Так, у мыши на этой стадии в каждой мейотической клетке образуется около 300 разрывов, а у лилии - несколько тысяч. В соматических клетках такое количество разрывов может появиться только в результате массированного облучения или обработки мощным мутагеном. Мейотическая клетка сама себе наносит эти повреждения с помощью белка Spo11 (гомолога топоизомеразы II архей). У архей нет мейоза, и этот белок участвует в репарации повреждений. Он разрезает поврежденную ДНК и воссоединяет свободные концы. У эукариот в мейозе белок Spo11 только режет, оставляя другим белкам заниматься воссоединением.
В воссоединении разорванных нитей ДНК активно участвует белок Rad51 (рис.5). У прокариот и в соматических клетках эукариот он задействован в репарации повреждений ДНК: в комплексе с другими белками связывается со свободными концами разорванных ДНК и внедряет их в ДНК гомологичных хромосом, одновременно расплетая ДНК-мишень. Задача внедренных участков состоит в том, чтобы найти комплементарные фрагменты определенной протяженности. К этому моменту гомологи уже прошли грубое выравнивание по штрих-коду, поэтому поиск происходит на относительно небольших расстояниях и именно в тех районах, где гомология наиболее вероятна. Найдя комплементарный участок, внедрившаяся нить ДНК спаривается с ним.

Рис.5. Мейотическая клетка обыкновенной бурозубки на стадии тонкого распознавания гомологов, окрашенная с использованием DAPI (специфичного для ДНК красителя), антител к рекомбинационному белку Rad51 и белку синаптонемного комплекса (SCP3) - осевого элемента мейотических хромосом.
Тонкое опознание заканчивается, когда количество связок между ДНК пары гомологичных хромосом достигает критического уровня. В мейотической клетке начинается разрезание связок. Большая часть разрезается и сшивается таким образом, что восстанавливается исходное состояние цепей ДНК (безобменный путь). Только небольшая их часть (у млекопитающих - меньше 0.1) сшивается крест-накрест (обменный путь), при этом ДНК одного из гомологов в пункте обмена соединяется с ДНК другого. Это и есть точки рекомбинации. Именно в них происходит переключение с одного гомолога на другой. В мейоз хромосомы вступили в таком виде, как они были получены от родителей, а выйдут из мейоза уже рекомбинантными. В материнской хромосоме будет часть отцовской, а в отцовской - часть материнской. Но это будут уже результаты рекомбинации, а мы пока вернемся к самому процессу.
Устранение опознавательных связок связано с нанесением новых повреждений в ДНК и репарацией этих повреждений. Эти процессы регулируются комплексом специфичных белков. Примечательно, что все они принадлежат к тем семействам белков, которые у прокариот и в соматических клетках эукариот участвуют в залечивании мутационных повреждений ДНК.
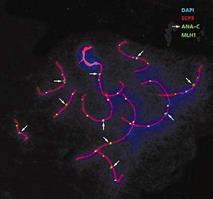
Рис.6. Мейотическая клетка обыкновенной бурозубки на стадии завершения рекомбинации, окрашенная с использованием DAPI, антител к белку репарации МLH1, центромерным белкам (ANA-C) и белку синаптонемного комплекса (SCP3).
Опознавательным знаком для точек рекомбинации служит белок MLH1 (рис.6). Он принадлежит к семейству белков мисматч репарации, функция которых - исправлять ошибки спаривания ДНК, т.е. устранять неспаренные нуклеотиды. Нас этот белок интересует в первую очередь как маркер точек рекомбинации. С помощью антител к MLH1, меченных флуоресцентными красителями, эти точки можно увидеть на хромосомах, а затем проанализировать частоту и распределение рекомбинационных событий по геному. Что мы и сделали на обыкновенной бурозубке. Для изучения мейоза она хороша тем, что у нее всего 10 пар хромосом, заметно отличающихся друг от друга по размеру и характеру дифференциального окрашивания [6]. Это позволило нам детально проанализировать особенности распределения точек рекомбинации по индивидуальным хромосомам.