Страница
3
модуляции изменчивости) (Laurençon et al., 1997). Молекулярные механизмы, участвующие в формировании этой системы еще не выяснены, однако наиболее вероятным представляется участие генов, которые одновременно контролируют процессы рекомбинации и репарации (Laurençon, Bregliano, 1995). Из известных на сегодняшний день генов в определении уровня реактивности наиболее вероятно участие генов mei-9+ и mei-41+ (Laurençon, Bregliano, 1995). Дальнейшее исследование роли, которую играет VAMOS в контроле генетической изменчивости при неблагоприятных условиях окружающей среды, может существенно прояснить работу молекулярных механизмов адаптации.
Дисгенные нарушения в рассмотренных системах гибридного дисгенеза в основном обусловлены транспозициями и эксцизиями мобильных элементов в развивающихся зародышевых клетках. Высокая частота хромосомных перестроек и рекомбинации у самцов происходят преимущественно в сайтах инсерции МГЭ (Engels, Preston, 1984; Sved et al., 1990). Повышенный уровень мутаций происходит от инсерционных мутаций и других индуцированных транспозициями МГЭ изменений в геноме.
Явление гонадной атрофии
Активация мобильных элементов в системах гибридного дисгенеза вызывает, среди прочих нарушений, особый вид стерильности гибридов, которая обусловлена недоразвитием гонад (Bregliano et al., 1980). Дисгенная стерильность по-разному проявляется в трех системах гибридного дисгенеза. P-M дисгенез приводит к недоразвитию яичников у гибридных самок и самцов (GD-стерильность) (рис. 1) (Kidwell et al., 1977; Schaeffer et al., 1979), в I-R системе, не происходит изменения морфологии гонад, но увеличивается уровень дефектных яиц и частота гибели эмбрионов (SF-стерильность) (Pelisson, 1979). Активация hobo элементов в H-E системе гибридного дисгенеза приводит как к недоразвитию гонад у самок и самцов первого поколения, так и к высокому уровню доминантных леталей среди отложенных яиц.
Стерильность является следствием потери зародышевых клеток на стадиях раннего эмбриогенеза и личинки (Niki, Chigusa, 1986). Для P-M гибридного дисгенеза гибель зародышевых клеток
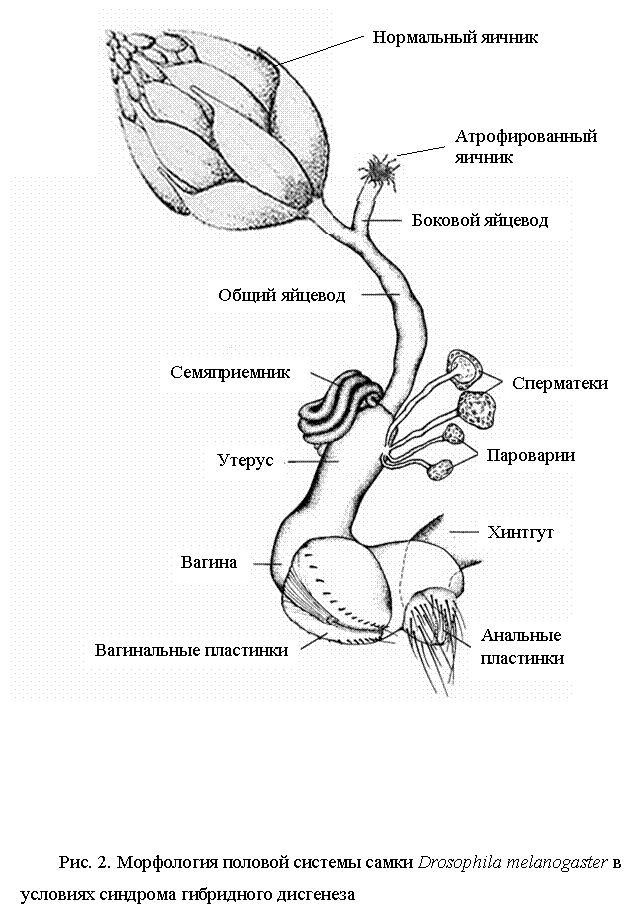 |
Атрофия гонад одно из характерных и интенсивно изучаемых аспектов P-M и H-E систем гибридного дисгенеза. GD-стерильность является следствием вымирания клеток в примордиальной зародышевой линии возможно из-за хромосомных разрывов, опосредованных активностью P-элемента (Niki, Chigusa, 1986). Однако ни кинетика гибели зародышевых клеток, ни число клеток, которые должны быть элиминированы для появления GD-стерильности не известно. Первые признаки стерильности появляются уже у 5-6 часовых эмбрионов. Максимальный уровень гибели клеток наблюдается на личиночной стадии развития, когда зародышевые клетки испытывают экспоненциальный рост, но некоторые умирающие клетки были обнаружены до деления клеток зародышевой линии (Niki, Chigusa, 1986). Механизм гибели клеток может быть обусловлен разрывами хромосом, имеющими летальный эффект. Следствием этого является отсутствие половых клеток в яичниках и семенниках и общее недоразвитие их у взрослых гибридных самок и самцов. Гибриды могут быть полностью стерильны, если редуцированы обе железы и частично фертильны, если атрофирован один семенник или яичник (рис.2) (Kidwell et al., 1977). Не атрофированные гонады у дисгенных гибридов часто отстают в своем развитии и содержат меньшее число яиц или сперматоцитов по сравнению с недисгенными особями (Ashburner, 1989). Степень выраженности гонадной атрофии сильно зависит от температуры, при которой идет развитие гибридов. В отношении Р-М системы атрофия наиболее значительна при 29° С у самок и при 27° С у самцов, а при 24° С и ниже практически отсутствует (Kidwell et al., 1977; Engels, Preston, 1984). Для Н-Е системы характерна наиболее сильная атрофия при 25° С и наименьшая при 29° С (Stamatis et al., 1989). При более низких температурах АГ наблюдается в меньшей степени. Количество и размерность копий Р- и hobo-элементов так же сильно влияют на способность линий индуцировать гибридный дисгенез, в связи с чем, частота гонадной атрофии может изменяться от нескольких до ста процентов (Yannopoulos et al., 1987; Kidwell et al., 1988; Rasmusson et al., 1993).
Активность мобильных элементов в P-M и H-E системах гибридного дисгенеза обусловливает мутабильность некоторых нестабильных локусов. В P-M системе наибольшую известность получил локус singed-weak (Engels, 1989), в H-E системе локус vgal (Bazin et al., 1993) и сконструированный маркерный элемент h(w+) (Calvi, Gelbart, 1994). Аллель snw обусловлена инсерцией двух дефектных P-элементов и делеция одного или другого из этих элементов в присутствии транспозазы полноразмерных элементов приводит к появлению соответствующих производных аллелей sne и sn(+).Повышенная мутабильность локуса vgal обусловлена активацией дефектных копий hobo в присутствии активных элементов этого семейства. Генетически сконструированный маркерный элемент h(w+) представляет hobo-элемент с встроенным геном mini-white, который определяет оранжевую окраску глаз (Calvi, Gelbart, 1994). Маркерная white линия несет в X-хромосоме два сконструированных элемента h(w+). Это обусловливает оранжевый фенотип цвета глаз, но при эксцизии одного из элементов цвет становится менее выраженным (Calvi, Gelbart, 1994). Линии с нестабильными аллелями широко используются при оценке активности P и hobo-элементов в дисгенных скрещиваниях, и частота мутирования этих аллелей служит дополнительным количественным критерием.
Таким образом, контроль активности мобильных генетических элементов в системах гибридного дисгенеза тесно взаимосвязан с механизмами транспозиций и репарации генетических повреждений. Это обусловливает чувствительность гибридного дисгенеза к действию внешних факторов и его модификацию различным генетическим фоном. Существующие предпосылки позволяют рассматривать синдром гибридного дисгенеза не только как показатель активности некоторых семейств мобильных генетических элементов, но и как целостную генетическую систему, обеспечивающую контроль генетической изменчивости генотипа в неблагоприятных условиях.