Страница
2
гибридов имеет значение также учет типа синапсиса отдельных хромосом и числа, образующихся хиазм. По числу бивалентов, унивалентов и мультивалентов, а также по типу синапсиса (плотный и рыхлый), числу хиазм бивалентов в мейозе судят о гомологии геномов и хромосом в наборе гибридного организма.
В полиплоидном ряду рода пшениц (Triticum) диплоидная пшеница-однозернянка Тг. monococcum имеет один геном (п = 7),
обычно обозначаемый А; тетраплоидные пшеницы (п = 14), например твердая пшеница (Tr. durum), имеют два разных генома А + В; гексаплоидные, например, мягкие пшеницы — три разных генома А + В + D. Таким образом, однозернянка оказывается диплоидом (АА = 2/г = 14), твердая пшеница — аллотетраплоидом (АА + + ВВ = 2/г = 14 + 14); мягкая пшеница — аллогексаплоидом (АА + ВВ + DD = 2/г = 14 + 14 + 14). Хромосомы генома А в мейозе, как правило, не конъюгируют с хромосомами геномов В и D.
Существует еще один вид аллотетраплоидной пшеницы Tr. Ti-mopheevi, открытый П. М. Жуковским в 1929 г., который имеет так же, как и твердая пшеница, 2n = 28 хромосом. Скрещивание этой пшеницы с твердой пшеницей осуществляется с трудом, и у гибридов сильно нарушен мейоз. В случае сходства геномов в мейозе должно было бы образовываться 14 бивалентов, но фактически наблюдается 7 бивалентов и 14 унивалентов (рис. 1). 7 хромосом у каждого из этих видов относятся к одному геному (АА), а 14 хромосом — к разным геномам. В случае скрещивания Tr. Timopheevi (2n = 28) с однозернянкой, имеющей геном А
(п = 7), в мейозе у гибрида обнаруживается менее 7 бивалентов и более 7 унивалентов. Следовательно, у Tr. Timopheevi есть геном А, сходный с таковым у однозернянки. Этот же геном выявляется по числу бивалентов и у твердой 28-хромосомной пшеницы. Второй геном Tr. Timopheevi, негомологичный также геному В твердой пшеницы, был обозначен G. Следовательно, у Tr. Timop-heevi есть два генома АА + GG. Как было установлено, род пшениц имеет 4 разных генома: А, В, D, G.
При скрещивании видов и цитологическом анализе степени конъюгации хромосом и образования бивалентов и унивалентов в мейозе удается выяснить гомологичность геномов и причины стерильности гибридов, вызванные несовместимостью геномов. Такой цитогенетический анализ геномного состава, по предложению Г. Кихары, с 1924 г. стали называть геномным анализом.
ПОЛУЧЕНИЕ ФЕРТИЛЬНЫХ ГИБРИДОВ
Получение плодовитых отдаленных гибридов достигается разными методами. При выборе метода имеют в виду цель восстановления плодовитости гибридов. Если необходимо сохранить в потомстве гибрида геномы обоих видов, то для этого пока известен только один способ, а именно применение амфидиплоидии. Плодовитые гибриды обычно получаются при этом только в тех случаях, когда межвидовая гибридизация приводит к хромосомной мутации, в результате которой образуется аллоплоид. Примером служит гибрид между капустой и редькой, полученный Г. Д. Карпеченко. Происходящие при такой гибридизации генетические изменения показаны на рисунке:
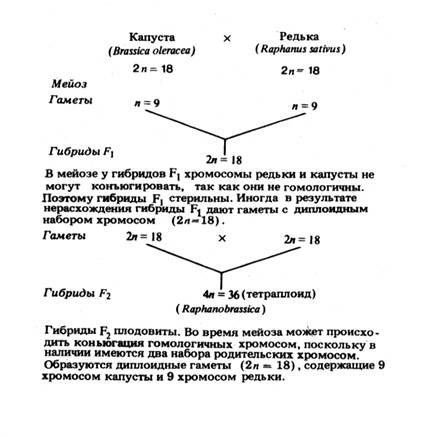
Предполагается, что первый отдаленный гибрид, возникший путем удвоения числа хромосом, был обнаружен в 1912 г. в результате скрещивания Primula floribunda (2n = 18) и P. verticillata (2/n =18). Один из гибридов от этого скрещивания, по-видимому, амфигаплоид, был стерильным, а другой — амфидиплоид (аллотетраплоид) — плодовитым и имел 2n = 36 хромосом. Гипотеза получения константных и плодовитых тетраплоидных форм у растений с помощью аллополиплоидии была предложена О. Винге еще в 1917 г.
В 1922—1924 гг. обстоятельное экспериментальное доказательство восстановления плодовитости путем аллополиплоидии было получено Г. Д. Карпеченко у межродового гибрида редьки и капусты.
В 1925 г. Дж. Клаусен и Т. Гудспид получили плодовитый гибрид от скрещивания Nicotiana tabacum (2n = 48) с N. glu-tinosa (2n = 24), имевшей 2n = 72 хромосомы. Первый межродовой гибрид пшеницы (2п = 42) и ржи (2n = 14) был получен в 1890 г. В. Римпау, но число хромосом у него не было установлено. Этот гибрид был назван Triticale.
В 1938 г. А. И. Державин при скрещивании твердой пшеницы (Тг. durum) с многолетней рожью (Secale montanum) впервые получил плодовитый не расщепляющийся амфидиплоид (аллогексаплоид) Triticale (2n = 42). Однако этот гибрид имел низкую плодовитость. Установлено, что у ржи есть геном S (или R), содержащий 7 хромосом, т. е. у диплоида — два генома SS, содержащие 2/г = 14. Геном S довольно успешно сочетается с двумя геномами твердой пшеницы (А и В) у гибрида Triticale, образуя аллогексаплоид AABBSS (2n = 42 хромосомы). Такие аллополиплоиды были получены также В. Е. Писаревым и др. Если рожь скрещивать с гексаплоидной мягкой пшеницей с геномами AABBDD (2n = = 42 хромосомы), то получаются октоплоидные Triticale с геномом AABBDDSS (2п = 56 хромосом). Однако все формы Triticale, как правило, имеют пониженную плодовитость.
Для повышения плодовитости каждую из этих гибридных форм скрещивали внутри себя (56 X 56 или 42 X 42 хромосом), однако заметного усиления плодовитости при этом не было достигнуто. Недавно венгерский генетик А. Киш произвел скрещивание между двумя формами Triticale: 56 X 42 и 42 X 56. В первом поколении растения были в значительной степени стерильны, но в следующих поколениях удалось отобрать формы с высокой урожайностью. Содержание глютеина и протеина в их зерне превышает таковое у ржи. Сейчас проводят испытание этих гибридов на песчаных почвах (рис. 2). При изучении мейоза у этих форм автор наблюдал поведение отдельного генома D и его потерю в том же самом скрещивании.

Рис. 2. Колосья фертильных линий Triticale, полученные А. Кишем.
Подобные аллополиплоиды могут быть истинными, когда у них сохраняются признаки обоих исходных видов и, следовательно, удвоенное число хромосом этих видов. Путем отбора удается повысить плодовитость за счет наилучшей сбалансированности геномов и нормализации мейоза. Однако следует иметь в виду, что при саморазмножении амфидиплоида геномы одного из исходных видов могут в ряду поколений элиминироваться, и тогда сохраняются геномы только одного вида (ложные амфидиплоиды). В этом случае повысится плодовитость, но такие особи не будут межвидовыми гибридами. В некоторых таких особях могут сохраняться лишь отдельные хромосомы утраченных геномов за счет частичной гомологии и включения их в геном одного из видов. Кроме того, возможно, что в результате транслокаций между негомологичными хромосомами некоторые локусы могут переместиться из одного генома в другой. В этом случае удвоение набора геномов скрещиваемых видов используют для восстановления парности гомологичных хромосом и плодовитости. Однако этот путь пригоден только для межвидовой и межродовой гибридизации растений.
Указанный способ применяют в селекции в том случае, когда преследуют цель получить от одного из скрещиваемых диплоидных видов ценные качества для создания новых сортов, а гибриды первого поколения оказываются стерильными. Этот метод был использован в ряде работ. Так, например, М. Ф. Терновский использовал аллополиплоидию в качестве моста, восстанавливающего плодовитость гибридов и передачу ценных свойств диких видов культурным формам табака. Обычно сорта табака не обладают устойчивостью к табачной мозаике и к мучнистой росе, в то время как некоторые дикие виды этими свойствами обладают. Вначале М. Ф. Терновский получил амфидиплоид (аллогексаплоид) от скрещивания Nicotiana glutinosa (n = 12) X N. tabacum (л = 24). В качестве мужского родителя был взят ценный стандартный сорт N. tabacum Дюбек Никитский 44. У амфидиплоида (аллогексаплоида) с 2п = 72 имелись парные геномы каждого вида, и мейоз протекал относительно нормально, лишь иногда наблюдалось образование унивалентов наряду с бивалентами. У этого гибрида проявлялась устойчивость к заболеванию табачной мозаикой, но качество табака значительно уступало Дюбеку Никитскому 44. Тогда данный аллогексаплоид вновь скрестили с Дюбеком Никитским 44, в результате чего был получен сесквидиплоид с одним геномом N. glutinosa и двумя геномами N. tabacum. В последующем этот частично плодовитый гибрид неоднократно скрещивался с табаком, и в потомстве вели отбор по продуктивности.