Цитоскелет сигнализирует
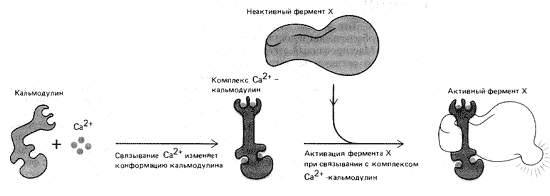
Рис. 12. Механизм активации Ca2+-зависимого ферментау
Присоединяющийся к кальмодулину кальций активирует его аналогично тому, как цАМФ стимулирует работу протеинкиназ. Именно так, например, происходит инициация сокращения гладкой мускулатуры. Высвободившиеся в ответ на внешний сигнал ионы кальция связываются с кальмодулином, который после этого взаимодействует с ферментом киназой и активирует ее (рис. 12). Комплекс киназа–кальмодулин связывается с актином, приводя его в рабочее состояние. В результате гладкие мышцы сокращаются. Опосредованный кальцием путь сигнала к поперечно-полосатой скелетной мускулатуре более длителен, зато гладкие мышцы в отличие от поперечно-полосатых могут значительно дольше находиться в сокращенном состоянии. Именно поэтому мускулы-замыкатели двустворчатых раковин могут часами сжимать свои створки.
В клетках разных тканей активированный кальцием кальмодулин связывается с различными белками-мишенями, влияя на их работу. Такое поведение кальмодулина вызывает вопрос, который возникал и при обсуждении влияния цАМФ и активируемой им протеинкиназы на активность белков, а именно: почему в разных клетках одни и те же активированные кальцием молекулы кальмодулина присоединяются к различным белкам?
Загадка стероидных гормонов
Совершенно аналогичная проблема возникает и при изучении гидрофобных стероидных гормонов, структура которых похожа на структуру жирорастворимого вещества холестерина. Термин «гидрофобный» указывает на их плохую растворимость в воде (от гр. hydor – вода и phobos – страх). Такие гормоны, будучи жирорастворимыми, легко проходят через состоящие из фосфолипидов клеточные мембраны. Оказавшись внутри клетки, стероидные гормоны связываются с соответствующими рецепторами. Рецепторы изменяют свою пространственную форму (конформацию) и, проникая в ядро через его ядерную мембрану, соединяются с определенными последовательностями нуклеотидов в ДНК, тем самым «включая» или «выключая» транскрипцию определенных генов. Такая последовательность событий доказана для стероидного гормона кортизона – его комплекс с рецептором связывается с соответствующим единственным геном, который удалось выделить и клонировать. Какая цепь превращений вызывается действием других гормонов и как происходит их специфическая «посадка» на определенный участок ДНК, во многом пока не ясно.
Не ясно и другое. Показано, что один и тот же гормон, связываясь со своим специфическим рецептором, вызывает различные ответы в разных клетках.
Забытое пространство
Число разнообразных сигналов, которые клетка может получать извне, очень велико. Это слабые электрические импульсы, гормоны, медиаторы, различные ростовые факторы и другие воздействия. Количество же вторичных мессенджеров, с помощью которых все это множество внешних сигналов влияет на внутриклеточные процессы, удивительно мало. Это цАМФ, ионы кальция, специальные молекулы типа высокофосфорилированных нуклеотидов (РРАРР – фосфат-фосфат-аденин-фосфат-фосфат) или инозитолтрифосфат.
Как же с помощью этих вторичных посредников клетка умудряется понять, какие именно сигналы их вызвали и каким образом необходимо на них реагировать? На этот вопрос трудно ответить прежде всего потому, что все клеточные компоненты (молекулы и ансамбли молекул) строго определенным образом скомпонованы в пространстве клетки.
Изучая по отдельности детали сложного часового механизма, который устроен несравнимо проще клетки, нелегко понять, как все эти шестеренки, маховички и пружины влияют друг на друга в работающем хронометре. Между тем именно такую задачу приходится решать исследователям клетки. Чтобы понять существо отдельных явлений или процессов, нужно разрушить клетку, выделить из нее белки, изучить их свойства и только потом попытаться установить их роль в том или ином процессе. При этом допускаются упрощения. Так, обычно принимается, что водорастворимые белки свободно диффундируют в цитоплазме наподобие крупинок в супе и никак не связаны между собой. Между тем само устройство некоторых клеточных органелл предполагает, что комплексы взаимодействующих ферментов должны образовывать специально сконструированные архитектурные ансамбли. Например, множество белков, катализирующих окислительные реакции в дыхательной цепи, располагаются на внутренних мембранах митохондрий в строго определенном порядке. Именно такая пространственная организация позволяет им с успехом осуществлять передачу богатых энергией электронов.
Но клетка содержит множество белков, связи которых друг с другом более лабильны и изменчивы во времени. По-видимому, для регуляции их пространственного взаимодействия требуется не прочное «заякоривание», а более тонкий и гибкий механизм. В частности, обеспечивать такую пространственную организацию могут белки цитоскелета. Они образуют настолько ажурные и динамичные структуры, что их изучение стало возможным лишь относительно недавно.
Строительные леса цитоскелета
Первые свидетельства сложности внутренней архитектуры цитоплазмы были получены еще в XIX в., когда в результате серебрения срезов тканей в клетках стали различать явственно проступающие сетеподобные структуры. Однако к изучению их состава и устройства удалось приступить лишь в 60-е гг. XX в., когда в биологии стали широко применяться такие тонкие методы исследований, как электронная микроскопия, ультрацентрифугирование и электрофорез.
В цитоплазме были обнаружены сложные структуры, образующие цитосклет. Выяснилось, что тяжи цитоскелета построены в основном из тонких (диаметром 7 нм) актиновых филаментов и длинных, толстых (диаметром 25 нм) и жестких микротрубочек, состоящих из - и -тубулина. Эти белки оказались очень лабильными, способными формировать легко изменяющиеся динамичные пространственные структуры. В частности, глобулярные белки актина не только легко и быстро полимеризуются в длинные вытянутые нити – филаменты (рис. 13). Они взаимодействуют с целым набором других вспомогательных белков, в результате чего возникает определенным образом организованная пространственная сеть филаментов.
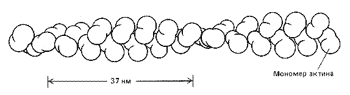
Рис. 13. Организация глобулярных молекул актина в актиновом филаменте
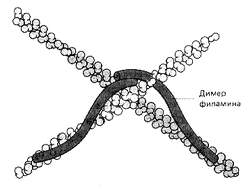
Рис. 14. Создание сети с помощью молекул филамина
Вспомогательные белки филамин и -актинин выполняют функции своеобразных скобок, сшивающих филаменты актина в структуру, напоминающую рыболовную сеть (рис. 14). Белок фибрин связывает актиновые филаменты в толстый пучок вроде веника или снопа. Тропомиозин стабилизирует уже сформированные тяжи актина (рис. 7). Гельзолин действует словно секатор, разрезая длинные филаменты на отдельные кусочки. Профилин, как нянька, сопровождает актиновые глобулы к местам их присоединения к образовавшимся ранее фрагментам нитей, виллин служит инициатором полимеризации актина в растворе, а тимозин, наоборот, не позволяет глобулярному актину соединятся в нити. Наконец, миозин способен активно подтягивать филаменты навстречу друг другу (рис. 5). Таким образом актиновые филаменты можно уподобить арматуре строительных лесов, которые можно резать, надставлять и соединять под любыми углами и в любых положениях. Их можно также соединять вместе, создавая тянущее напряжение всей конструкции.