Страница
2
Аппликация миансерина снижала амплитуду вызванных сокращений. Статистически значимый эффект миансерина проявлялся в концентрациях от 10-4 до 10-3 М (рис. 2а). Более низкие концентрации миансерина (5×10-5 М и ниже) статистически выявляемого влияния на амплитуду мышечных сокращений не оказывали. Эффект действия миансерина, в отличие от серотонина, проявлялся непосредственно после его аппликации.
Аппликация метерголина также снижала амплитуду вызванных мышечных сокращений. Однако статистически значимый эффект метерголина проявлялся в концентрациях от 10-5 до 10-4 М (рис. 2б). Эффект действия метерголина проявлялся спустя 15 минут после его аппликации.
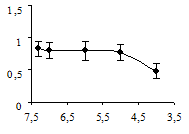
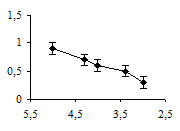
а б
Рис. 2. Концентрационные зависимости влияния миансерина (а) и метерголина (б) на сокращения дорсальной продольной мышцы Lymnaea stagnalis L., вызванные ритмической стимуляцией n. cervicalis inferior: оси абсцисс – отрицательный десятичный логарифм концентраций исследованных препаратов в нормальном физиологическом растворе; оси ординат – отношение амплитуды мышечных сокращений в физиологическом растворе к амплитуде мышечных сокращений в метерголине или миансерине; вертикальные линии – стандартное отклонение (n = 5)
Обсуждение
Усиливающее действие серотонина (5-HT) на нервно-мышечную передачу было отмечено у многих видов моллюсков [3 – 5]. Известно, что действие этого вещества проявляется через большое количество типов и субтипов рецепторов. К настоящему времени у позвоночных животных было клонировано семь различных 5-HT рецепторов: 5-HT1A, 5-HT1B, 5-HT1C, 5-HT1Da, 5-HT1Db, 5-HT2 и фармакологически не охарактеризованный 5-HT рецептор человека [16]. Однако сведения о 5-HT рецепторах в нервной системе беспозвоночных в большинстве случаев отрывочны и не поддаются классификации, установленной для позвоночных животных.
Как показали наши наблюдения, серотонин усиливает, а его антагонисты – миансерин и метерголин – подавляют амплитуду вызванных сокращений дорсальной продольной мышцы. Возможно, что механизм увеличения амплитуды вызванных мышечных сокращений в присутствии серотонина может указывать на возможную роль последнего как медиатора нервно-мышечной передачи. В этом случае взаимодействие экзогенного серотонина с синаптическими 5-HT рецепторами мышечных клеток может приводить к их деполяризации и повышению возбудимости. Частичное сокращение исследуемой мышцы при добавлении серотонина в физиологический раствор, вероятно, согласуется с таким предположением. Исчезновение усиливающего действия экзогенного серотонина при повышении его концентрации до 10-6 М мы склонны объяснять развивающейся десенситизацией 5-HT рецепторов. Угнетающее действие антагонистов серотонина на мышечные сокращения также может быть следствием их взаимодействия с синаптическими рецепторами. В наших опытах метерголин и миансерин проявляли свое действие в значительно более высоких концентрациях по сравнению с серотонином. В таких концентрациях эти вещества могут блокировать одновременно 5-HT1A и 5-HT2 типы серотониновых рецепторов, обнаруженные у L. stagnalis, фармакологический профиль которых в исследованных гигантских церебральных клетках совпадает с профилем этих рецепторов у позвоночных [17 – 19]. Поэтому влияние низких концентраций серотонина можно было бы объяснить взаимодействием его молекул с другим типом серотониновых рецепторов, например 5-HT3. Последний был обнаружен в нервной системе L. stagnalis при исследовании фармакологических свойств нейронов церебрального ганглия [20].
К настоящему моменту известно, что набор агонистов и антагонистов для 5-HT рецепторов беспозвоночных значительно отличается от набора фармакологических агентов 5-HT рецепторов позвоночных животных. К примеру, константа диссоциации (Ki, нМ) для метерголина в отношении 5-HTlym составляет 55, а при связывании с 5-HT1 и 5-HT2 рецепторами позвоночных животных 10 и 2,1 соответственно [21, 16]. Отмечается, что 5-HT рецепторы беспозвоночных животных характеризуются иными фармакологическими и функциональными свойствами, чем рецепторы позвоночных [22]. Кроме того, филогенетические различия между моллюсками и позвоночными животными настолько велики, что фармакологический профиль соответствующих рецепторов беспозвоночных видов вряд ли может быть полностью подобен профилю рецепторов позвоночных животных, поэтому требует для дифференциации других фармакологических агентов [23, 24]. Возможно, что в данном нервно-мышечном соединении присутствует особый тип серотониновых рецепторов, чей фармакологический профиль отличается от уже известных. Высокие же значения эффективных концентраций мианзерина и метерголина могут свидетельствовать о высоком значении константы диссоциации этих веществ для данного типа рецептора. По крайней мере, фармакологическая характеристика 5-HTLym рецептора, недавно полученного методом клонирования в культуре нервных клеток L. stagnalis, не совпадает ни с одним из фармакологических профилей 5-HT1 рецепторов позвоночных. На основе чего авторы сделали предположение о дополнительном субклассе – 5-HT1 – подобных рецепторов, еще не обнаруженном у позвоночных [16]. Для поиска ответа на вопрос: сходны ли свойства серотониновых рецепторов нервно-мышечного соединения и центральных нейронов моллюска L. stagnalis – необходимо существенно расширить спектр исследуемых соединений.
Авторы приносят благодарность профессорам Ф.-В. Шурману (университет Геттингена) и Г. Брилла за любезно представленные фармакологические препараты, а также К.А. Судоплатову за техническую помощь в организации экспериментов. Работа выполнена при финансовой поддержке грантов ИНТАС-РФФИ №97-04-71075 (IR-97-798) и Минобразования №97-0-10.0-207.
Списоклитературы
1. Muneoka Y., Twarog B. Neuromuscular transmission and excitation-contraction coupling in molluscan muscle // The Mollusca / Eds. A.S.M. Saleuddin, K.M. Wilbur. New-York; London: Academic Press, 1983. V. 4. P. 35 – 64.
2. Heyer C.B., Katert S.B., Karlsson U.L. Neuromuscular systems in mollusca // Amer. Zool. 1973. V. 13. P. 217 – 270.
3. Fox L.E., Lloyd P.E. Serotonergic neurons differentially modulate the efficacy of two motor neurons innervating the same muscle fibers in Aplysia // J. Neurophysiology. 1998. V.80. P. 647 – 655.
4. Versen B., Gokorsch S., Fiedler A., Schipp R. Monoamines and the isolated auricle of Sepia officinalis: are there b-like receptors in the heart of a cephalopod? // J. Exp. Biol. 1999. V. 202. P. 1067 – 1079.
5. Yoshida M., Kobayashi M. Modulation of the buccal muscle contraction by identified serotonergic and peptidergic neurons in the snail Achatina fulica // J. Exp. Biol. 1995. V. 198. 729 – 738.
6. Fox L.E., Lloyd P.E. Glutamate is a fast excitatory transmitter at some buccal neuromuscular synapses in Aplysia // J. Neurophysiology. 1999. V. 82. P. 1477 – 1488.
7. Cook A. The withdrawal response of a freshwater snail (Lymnaea stagnalis) // J. Exp. Biol. 1975. V.62. P. 783 – 796.