Фармакологические свидетельства присутствия 5-НТ рецепторов в нервно-мышечном соединении брюхоногих моллюсков
Исследовано влияние аппликации серотонина и его двух антагонистов (миансерина и метерголина) на сокращения дорсальной продольной мышцы гастроподного моллюска Lymnaea staganalis L., вызванные электрическим раздражением n. cervicalis inferior. Установлено, что увеличение концентрации серотонина в физиологическом растворе вызывало двоякое действие: в концентрации 2×10 -8 – 10 -6 М он усиливал мышечные сокращения, а в концентрациях свыше 10 -5, напротив, ослаблял. Блокирующее действие антагонистов серотонина на амплитуду вызванных сокращений возрастало с увеличением их концентраций в исследованном диапазоне от 10 -5 – 10 -3 М (для миансерина) и от 5×10 ‑8 – 10 -4 М (для метерголина). Предполагается, что 5-HT рецепторы, участвующие в нервно-мышечной передаче в исследованном соединении, имеют иной фармакологический профиль, чем 5-HТ рецепторы позвоночных животных.
Введение
Основной объем сведений о механизмах нервно-мышечной передачи дают исследования синапсов на мускулатуре позвоночных и членистоногих животных. Соответствующие эксперименты, проведенные на брюхоногих моллюсках, сравнительно немногочисленны. И это притом, что разнообразие как гистологии и свойств мышечных волокон, так и их функциональных взаимоотношений с нейронами у брюхоногих может быть не меньше, чем у животных других систематических групп. Так же, как и у членистоногих, каждая мышечная клетка этих животных может быть иннервирована функционально различными типами волокон – как возбуждающими (фазными или тоническими), так и тормозными [1]. Сами мышечные волокна могут быть связаны между собой электрически, что в большинстве случаев оставляет открытым вопрос о числе и типе иннервирующих их аксонов, которые могут образовывать на миоцитах многочисленные окончания [2]. Столь же разнообразной представляется химическая природа медиаторов нервно-мышечной передачи у этих животных. Наиболее распространенными типами нейротрансмиттеров являются соединения группы моноаминов (ацетилхолин, серотонин, дофамин), а также аминокислоты (глутамин) и нейропептиды (FMRF-амид) [3 – 6]. Поэтому любые исследования, дополняющие пока еще сравнительно разрозненную картину сведений о нервно-мышечной передаче брюхоногих моллюсков, важны для эволюционного анализа физиологических функций животного организма.
В качестве модели исследования было выбрано нервно-мышечное соединение n. cervicalis inferior с продольной дорсальной мышцей Lymnaea stagnalis, которая является одним из эффекторов оборонительного рефлекса [7]. Известно, что тела управляющих этой мышцей мотонейронов обнаружены в большинстве ганглиев ЦНС моллюска [8]. Эти нейроны принимают возбуждающие сигналы с механо- и фоторецепторов кожной поверхности тела и направляют свои аксоны к указанной мышце в составе n. cervicalis inferior и n. cervicalis superior [9; 10]. Сведения о структурной организации синапсов, присутствующих на миоцитах продольной дорсальной мышцы Lymnaea stagnalis в литературе отсутствуют. Однако предыдущими исследованиями было показано угнетающее действие двухвалентных катионов на амплитуду вызванных мышечных сокращений, что может свидетельствовать о химическом механизме нервно-мышечной передачи [11]. Одновременно было показано участие в этом процессе серотонинергических механизмов и соответствующих рецепторов. Настоящая работа представляет собой шаг в направлении детализации их фармакологических свойств этих механизмов.
Материалы и методы
Животные. Опыты выполнены на взрослых особях Lymnaea stagnalis (высота раковины около 3 см), собранных в прудах Калининграда, которых содержали в лабораторных условиях.
Препарат состоял из ноги и мантии моллюска, рассеченной по средней линии. Центральный конец нерва, n. cervicalis inferior (номенклатура по [12]), иннервирующего дорсальную продольную мышцу (название мышцы дается по [13]), помещали во всасывающий электрод.
Состав физиологического раствора (мМ): NaCl – 40; KCl – 3; CaCl2 – 3; MgCl2 – 1. Значение РН = 7,5 – 7,6 поддерживали карбонатным буфером (NaHCO3).
Исследуемые вещества. В ходе экспериментов применяли серотонин (5-НТ), миансерин и метерголин (Sigma Chemical Co). Маточные растворы веществ готовили на дистиллированной воде (серотонин, миансерин) или на 0,1 Н растворе HCl (метерголин). Рабочие растворы получали путем разведения маточных в физиологическом.
Оборудование. Датчик для регистрации сокращения мышцы был изготовлен из пары проволочных тензосопротивлений, наклеенных на противоположные стороны полоски гибкой пленки [14]. Сопротивление тензодатчика изменялось при деформации пленки, вызванной натяжением лески, прикрепленной с помощью металлического крючка к исследуемой мышце. Исходное натяжение лески устанавливали вручную винтом манипулятора. Тензосопротивления включали в схему мостика Уитстона, с которого электрический сигнал подавали на вход усилителя постоянного тока. Усиленный сигнал поступал на осциллограф С1-67 и чернильный автоматический потенциометр К-201 (Германия).
Стимуляцию нерва осуществляли сериями электрических импульсов (длительностью 2 мс, амплитудой около 5 В), которые подавали на хлорсеребряный электрод, вмонтированный в трубочку всасывающего электрода. Второй хлорсеребряный электрод помещали в физиологический раствор в экспериментальной камере.
Ход эксперимента
1. Электрическая стимуляция препарата в физиологическом растворе.
2. Замена физиологического раствора на растворы веществ и спустя 10 минут электрическая стимуляция нерва. Перед каждым новым раствором с последовательно увеличивающейся концентрацией вещества препарат промывали в физиологическом растворе в течение 30 минут.
3. Промывка препарата в физиологическом растворе в течение 30 минут и электрическая стимуляция.
Статистическая оценка полученных результатов проводилась по Т-критерию Уилкоксона [15].
Результаты
Аппликация серотонина в концентрациях в диапазоне от 2×10‑8 М до 10-6 М усиливала мышечные сокращения, причем эффект наблюдался спустя 10 минут после добавления вещества в ванночку и мог сохраняться на протяжении получаса после возврата к физиологическому раствору. Более высокие концентрации серотонина (10-5 М и выше), напротив, снижали амплитуду мышечных сокращений. Максимальный усиливающий эффект серотонина проявлялся при его концентрации в ванночке около 10-7 М (рис. 1). Эта концентрация серотонина сама по себе вызывала небольшое укорочение мышцы. Изменения амплитуды сокращений, которые наблюдались в присутствии серотонина 10-8 и 10-6 М, статистически не подтверждались.
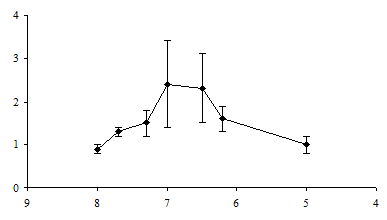
Рис. 1. Концентрационная зависимость влияния серотонина на сокращения дорсальной продольной мышцы Lymnaea stagnalis L., вызванные ритмической стимуляцией n. cervicalis inferior:
ось абсцисс – отрицательный десятичный логарифм концентрации серотонина в нормальном физиологическом растворе; ось ординат – отношение амплитуды мышечных сокращений в растворе серотонина к амплитуде мышечных сокращений в физиологическом растворе; вертикальные линии – стандартное отклонение (n = 5)