Страница
3
Избыточная продукция нейронов, после которой следует период гибели клеток, является характерной чертой развития нервной системы позвоночных. Некоторые из нейронов, которые погибают, возможно, не смогли образовать синапсов, или образовали их с неподходящей клеткой-мишенью. В подобных случаях гибель клеток связана с особенностью иннервации. Однако большинство клеток, которые гибнут, достигают и иннервируют правильные клетки-мишени. Таким образом, гибель клеток является основным механизмом, который поддерживает равновесие между количеством нейронов и их мишеней.
Неожиданной находкой явилось то, что ингибиторы образования мРНК или синтеза белка предотвращали гибель нейронов в отсутствие необходимых им нейротрофинов. Результаты этих и более поздних экспериментов показали, что гибель нейронов типично происходит путем апоптоза. Апоптоз — это процесс, который активирует внутреннюю «суицидальную» программу клетки, которая приводит к упорядоченному (запрограммированному) распаду ДНК и белков в клетке. Существенно, что для этого необходим предварительный синтез протеолитических ферментов или их активаторов.
Уменьшение числа связей и исчезновение полинейрональной иннервации
После того как популяция нейронов, иннервирующих определенную мишень, уменьшается благодаря гибели клеток, оставшиеся нейроны конкурируют друг с другом за синаптическую территорию. Это конкурирование обычно приводит к потере некоторых первично образованных веточек и синапсов («обрезка»). Уменьшение числа связей обеспечивает механизм для формирования правильной и полной иннервации мишени определенной популяцией нейронов. В некоторых случаях механизм уменьшения числа связей также обеспечивает механизмы коррекции ошибок; в других случаях он отражает стратегию поиска пути аксоном.
Особо яркий пример конкурентного уменьшения числа связей можно увидеть в развивающейся скелетной мышце. У взрослых животных каждый мотонейрон иннервирует группу до 300 мышечных волокон, формируя двигательную единицу , но каждое мышечное волокно иннервируется только одним аксоном. Однако в развивающейся мышце мотонейроны бурно ветвятся, так что каждое мышечное волокно в итоге иннервируется аксонами от нескольких мотонейронов (рис. 4), что называется полинейрональной иннервацией. В каждом развивающемся мышечном волокне синаптические окончания всех аксонов разбросаны вокруг одного участка, вблизи скоплений рецепторов АХ и других компонентов постсинаптического аппарата. По мере развития веточки аксонов исчезают, что приводит в конечном итоге к формированию взрослого паттерна. Этот процесс не вызывает гибель клетки (которая обычно происходит на более ранних стадиях развития), а только уменьшение количества мышечных волокон, иннервируемых каждым из мотонейронов.
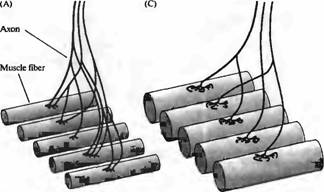
Рис. 4. (Рис. 4В, 4D см. на цветной вклейке после с. 640.) Полинейрональная иннервация и ее исчезновение в нерв но мышечном соединении позвоночных. (А) Во время развития эмбриона веточки аксонов моторных нейронов иннервируют множество мышечных волокон, и каждое мышечное волокно иннервируется несколькими мотонейронами (полинейрональная иннервация). (В) Флуоресцентная микрограмма нервно-мышечного соединения мыши (стадия Е18), показывающая распределение терминалей двух аксонов, каждый из которых помечен при помощи липофильной метки. В период полинейрональной иннервации терминальные ветвления аксонов мотонейронов, иннервирующих определенное мышечное волокно, переплетаются в областях отдельных синапсов. (С) После рождения полинейрональная иннервация исчезает по мере того, как веточки аксонов подвергаются ретракции, в результате чего каждое мышечное волокно получает иннервацию только от одного мотонейрона. (D) Флуоресцентная микрофотография нервно-мышечного соединения мыши во время исчезновения полинейрональной иннервации. Два аксона, иннервирующих синапс, были помечены как в В. Все терминали одного аксона исчезли, и сам аксон подвергся ретракции.
Исчезновение полинейронной иннервации регулируется конкуренцией между аксонами различных мотонейронов за синаптическое пространство на мышечных клетках. Наиболее ярким примером являются эксперименты на развивающихся мышцах лапки крысы. Когда все кроме одного моторные аксоны, иннервирующие эту мышцу, были перерезаны на раннем этапе развития, оставшийся аксон подвергся ветвлению и начал иннервировать большое количество волокон данной мышцы. В то время, когда обычно происходит исчезновение полинейрональной иннервации, ни один синапс не исчез. В отсутствие конкуренции для выживания мотонейрон поддерживает контакты с каждой миофибриллой, которую он ранее иннервировал. В экспериментах Лихтмана с коллегами были получены яркие картинки этого процесса при помощи визуализирования нервных терминалеи у животных in vivo при помоши витальных красителей и наблюдения за изменениями синаптической структуры во время исчезновения синапсов. Подобная же ретракция избыточного количества связей была показана в автономном ганглии неонатальных крыс и морских свинок. Каждая ганглионарная клетка первоначально получает большое количество связей, больше пяти, однако к возрасту 5 недель из них обычно остается только одна.
Активность нервов и исчезновение синапсов
Физиологические эксперименты показывают, что активность нейронов играет определенную роль в исчезновении синапсов, влияя как на интенсивность, так и на и результат конкуренции между терминалями аксонов. Стимулирование мышцы через нерв при помощи имплантированных металлических электродов увеличивает интенсивность исчезновения синапсов. Уменьшение активности путем добавления тетродотоксина в пространство вокруг нерва для блокирования потенциалов действия приводит к ингибированию синаптической передачи и замедлению исчезновения синапсов. На мышцах, которые получают сигналы от аксонов двух различных нервов, возможно проведение интересных экспериментов по блокированию проведения импульсов избирательно в одном из нервов. В таких случаях неактивные нейроны не могут, очевидно, нормально конкурировать: аксоны блокированных нейронов иннервируют двигательных единиц меньше, чем в норме; аксоны активных нервов иннервируют волокон больше, чем обычно. Однако доминирование неблокированных нервных волокон не полное, что говорит о роли других факторов, кроме активности, в исчезновении синапсов.
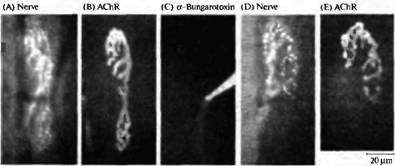
Рис. 5. Конкуренция между веточками одного мотонейрона, зависящая от активности. (А, В) Флуоресцентные микрофотографии нервно-мышечного соединения грудинососцевидной мышцы мыши, показывающие пресинаптическую терминаль (А), окрашенную при помощи 4-Di-2-Asp (который окрашивает митохондрии) и постсинаптическую мембрану (В), окрашенную при помощи родамин-связанного бунгаротоксина в низкой дозе (который окрашивает рецепторы АХ). (С) Рецепторы АХ в нижней части соединения блокированы при помощи насыщающей дозы немеченного -бунгаротоксина, что приводит к блокаде нервно-мышечной передачи селективно в этой области синапса. (D, Е) Через 31 день тот же самый синапс, что показан на А и В, был заново исследован. Терминаль аксона (D) и постсинаптические рецепторы (Е) исчезли из блокированной области. Таким образом локальная блокада передачи приводит к локальному исчезновению синапса.