Структурно-функциональная организация генетического материала
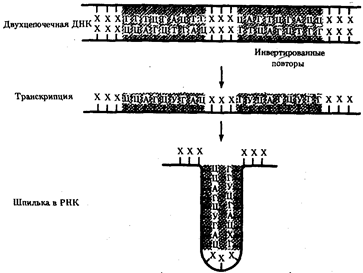
Рис.38. Образование шпильки участком РНК при терминации транскрипции у прокариот
Область РНК, несущая палиндром, образует комплементарно спаривающуюся структуру - шпильку (инвертированные повторы заштрихованы)
В полицистронной мРНК прокариот между кодирующими участками имеются межцистронные области, варьирующие по размерам (рис.3.39).
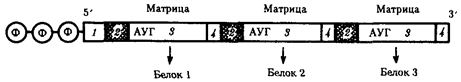
Рис.39. Полицистронная матричная РНК прокариот:
1 - некодирующие области, 2 - межцистронные области, 3 - кодирующие области, 4 - терминирующие кодоны
В связи с тем что прокариотические гены целиком состоят из нуклеотидных последовательностей, участвующих в кодировании информации, транскрибированные с них РНК сразу после их синтеза способны выполнять функцию матриц для трансляции. Лишь в исключительных случаях требуется их предварительное созревание - процессинг.
В отличие от прокариотических генов большинство генов эукариотических клеток прерывисты, так как несут в своем составе неинформативные нуклеотидные последовательности - интроны, не участвующие в кодировании информации. В связи с этим первичные транскрипты, синтезированные РНК-полимеразой II, обладают большими, чем необходимо для трансляции, размерами и оказываются менее стабильными. В совокупности они образуют так называемую гетерогенную ядерную РНК (тРНК), которая прежде чем выйти из ядра и начать активно функционировать в цитоплазме, подвергается процессингу и превращается в зрелые мРНК.
Процессинг эукариотических мРНК. Созревание, или процессинг, мРНК предполагает модифицирование первичного транскрипта и удаление из него некодирующих интронных участков с последующим соединением (сплайсингом) кодирующих последовательностей - экзонов. Модифицирование первичного транскрипта эукариотической мРНК начинается вскоре после синтеза его 5'-конца, содержащего одно из пуриновых оснований (аденин или гуанин). На этом конце образуется колпачок - кэп, который блокирует 5'-конец мРНК путем присоединения к первому нуклеотиду транскрипта трифосфонуклеозида, содержащего гуанин, связью 5'-5'.
Гффф + фффАфN… → ГфффАфN. + фф + ф В результате образуется последовательность ГфффАфЧМ ., в которой остаток туанина находится в обратной ориентации по отношению к другим нуклеотидам мРНК. Модификация 5'-конца мРНК предполагает также метилирование присоединенного гуанина и первых двух-трех оснований первичного транскрипта (рис.3.40). Образуемые на 5' - концах мРНК кэпы обеспечивают узнавание молекул мРНК малыми субчастицами рибосом в цитоплазме. Кэширование осуществляется еще до окончания синтеза первичного транскрипта.
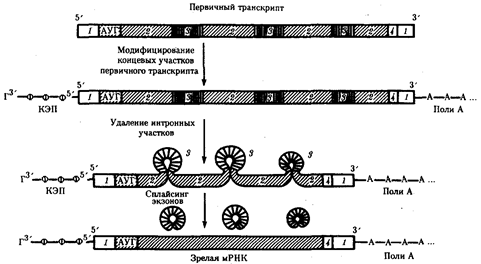
Рис. 40. Образование зрелой мРНК эукариот в ходе процессинга:
1 - некодирующие последовательности, 2 - экзоны, 3 - интроны, 4 - кодон-терминатор
После завершения транскрипции происходит удаление части нуклеотидов на 3'-конце первичного транскрипта и присоединение к нему последовательности, состоящей из 100-200 остатков адениловой кислоты (полиА) (рис.3.40). Считают, что эта последовательность способствует дальнейшему процессингу и транспорту зрелой мРНК из ядра. После выхода мРНК в цитоплазму ее полиА-последовательность постепенно укорачивается под действием ферментов, отщепляющих нуклеотиды на 3'-конце. Таким образом, по длине полиА-последовательности можно косвенно судить о времени пребывания мРНК в цитоплазме. Возможно, добавление полиА-последовательности в ходе процессинга повышает стабильность мРНК. Однако около трети мРНК вообще не содержат полиА-участка. К ним относятся, например, гистоновые мРНК.
Образование кэпа на 5'-конце и полиА-последовательности на 3'-конце характерно только для процессинга РНК, синтезируемых РНК-полимеразой II. Кроме метилирования при формировании кэпов в мРНК высших эукариот происходит метилирование небольшой части внутренних нуклеотидов с частотой приблизительно одно на тысячу оснований мРНК.
Наряду с модифицированием мРНК эукариот процессинг предполагает удаление из первичных транскриптов неинформативных для данного белка интронных участков, размер которых варьирует от 100 до 10 000 нуклеотидов и более. На долю интронов приходится около 80% всей гяРНК. Удаление интронов с последующим соединением экзонных участков называют сплайсингом (рис.40).
Сплайсинг представляет собой механизм, который должен обеспечивать удаление из первичного транскрипта строго определенных интронных участков. Нарушение этого процесса может привести к сдвигу рамки считывания при трансляции и невозможности синтеза нормального пептида. Закономерность вырезания интронов, очевидно, обеспечивается благодаря наличию на их концах специфических нуклеотидных последовательностей, служащих сигналами для сплайсинга.
В настоящее время описано несколько вероятных механизмов сплайсинга, обеспечивающих точность этого процесса. Возможно, она достигается действием каких-то ферментов, специфически узнающих концевые участки интронов и катализирующих разрыв фосфодиэфирных связей на границе экзон - интрон, а затем образование связей между двумя экзонами.
Установлено активное участие в сплайсинге особых малых, ядерных РНК (мяРНК), образующих комплексы с белками (мяРНП). Очевидно, мяРНК своими нуклеотидными последовательностями комплементарно взаимодействуют с концевыми участками интронов, которые образуют при этом замкнутые петли. Расщепление РНК в устье интронной петли приводит к удалению неинформативной последовательности и соединению (сплайсингу) сближенных концов экзонов.
Обсуждается также автокаталитическая способность РНК-транскрипта к сплайсингу. Описанные способы сплайсинга свидетельствуют об отсутствии универсального механизма этого процесса, однако во всех случаях достигается точное удаление интронов с образованием определенной мРНК, обеспечивающей синтез необходимого клетке белка.
В настоящее время доказана возможность альтернативного (взаимоисключающего) сплайсинга, при котором из одного и того же первичного транскрипта могут удаляться разные нуклеотидные последовательности и образовываться разные зрелые мРНК. В результате одна и та же последовательность нуклеотидов ДНК может служить информацией для синтеза разных пептидов. Альтернативный сплайсинг, вероятно, очень характерен в системе генов иммуноглобулинов у млекопитающих, где он позволяет формировать на основе одного транскрипта мРНК для синтеза разных видов антител.
Благодаря преобразованиям, происходящим с РНК-транскриптом в ходе процессинга, зрелые мРНК эукариот характеризуются большей стабильностью по сравнению с прокариотическими мРНК.
По завершении процессинга зрелая мРНК проходит отбор перед выходом в цитоплазму, куда попадает всего 5% гяРНК. Остальная часть расщепляется, не покидая ядра.
Таким образом, преобразования первичных транскриптов эукариотических генов, обусловленные их экзонитронной организацией и необходимостью перехода мРНК из ядра в цитоплазму, определяют особенности реализации генетической информации в эукариотической клетке.