Строение и физиологические особенности рыб
Для поддержания оптимальной концентрации кислорода в воде, обеспечивающей наиболее эффективное течение физиологических процессов в организме рыб, нужно использовать аэрационные установки.
К небольшому пересыщению кислорода рыбы адаптируются быстро. У них повышается обмен и как результат увеличивается потребление корма и снижается кормовой коэффициент, развитие эмбрионов ускоряется, отходы снижаются.
Для нормального дыхания рыб очень важно содержание в воде СО2. При большом количестве двуокиси углерода дыхание рыб затруднено, так как уменьшается способность гемоглобина крови связывать кислород, насыщение кислородом крови резко снижается и рыба задыхается. При содержании CO2 в атмосфере 1 .5% СО2; крови не может поступать наружу, а кровь не может принимать кислород даже из насыщенной кислородом воды.
Кровеносная система
Главным отличием кровеносной системы рыб от других позвоночных является наличие одного круга кровообращения и двухкамерного сердца, наполненного венозной кровью (за исключением двоякодышащих и кистеперых).
Сердце состоит из одного желудочка и одного предсердия и помещается в околосердечной сумке, сразу за головой, позади последних жаберных дуг, т. е. по сравнению с другими позвоночными сдвинуто вперед. Перед предсердием имеется венозная пазуха, или венозный синус, со спадающими стенками; через эту пазуху кровь поступает в предсердие, а из него — в желудочек.
Расширенный начальный участок брюшной аорты у низших рыб (акулы, скаты, осетровые” двоякодышащи╦) образует сокращающийся артериальный конус, а у высших рыб — луковицу аорты, стенки которой сокращаться не могут. Обратному току крови препятствуют клапаны.
Схема кровообращения в самом общем виде представлена следующим образом. Венозная кровь, заполняющая сердце, при сокращениях сильного мускульного желудочка через артериальную луковицу по брюшной аорте направляется вперед и поднимается в жабры по приносящим жаберным артериям. У костистых рыб их четыре с каждой стороны головы—по числу жаберных дуг. В жаберных лепестках кровь проходит через капилляры и окисленная, обогащенная кислородом направляется по выносящим сосудам (их также четыре пары) в корни спинной аорты, которые затем сливаются в спинную аорту, идущую вдоль тела назад, под позвоночником. Соединение корней аорты спереди образует характерный для костистых рыб - головной круг. Вперед от корней аорты ответвляются сонные артерии.
От спинной аорты идут артерии к внутренним органам и мускулатуре. В хвостовом отделе аорта переходит в хвостовую артерию. Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь венозные капилляры впадают в вену, несущую кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (яремными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуются два Кювьерова протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта (желудка, кишечника) и селезенки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус
Рис. 1 Схема кровеносной системы костистой рыбы:
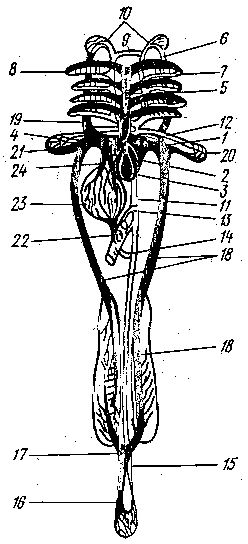 1 - венозная пазуха; 2 - предсердие; 3 - желудочек; 4 - луковица аорты; 5 - брюшная аорта; 6 - приносящие жаберные артерии; выносящие жаберные артерии; 8 - корни спинной аорты; 9 - передняя перемычка, соединяющая корни аорты; 10 - сонная артерия; 11 - спинная аорта; 12 - подключичная артерия; 13 - кишечная артерия; 14 - брыжеечная артерия; 15 - хвостовая артерия; 16 - хвостовая вена; 17 - воротные вены почек; 18 - задняя кардинальная вена; 19 - передняя кардинальная вена; 20 - подключичная вена; 21 - Кювьеров проток; 22 - воротная вена печени; 23 - печень; 24 - печеночная вена; черным показаны сосуды с венозной кровью, белым с артериальной.
1 - венозная пазуха; 2 - предсердие; 3 - желудочек; 4 - луковица аорты; 5 - брюшная аорта; 6 - приносящие жаберные артерии; выносящие жаберные артерии; 8 - корни спинной аорты; 9 - передняя перемычка, соединяющая корни аорты; 10 - сонная артерия; 11 - спинная аорта; 12 - подключичная артерия; 13 - кишечная артерия; 14 - брыжеечная артерия; 15 - хвостовая артерия; 16 - хвостовая вена; 17 - воротные вены почек; 18 - задняя кардинальная вена; 19 - передняя кардинальная вена; 20 - подключичная вена; 21 - Кювьеров проток; 22 - воротная вена печени; 23 - печень; 24 - печеночная вена; черным показаны сосуды с венозной кровью, белым с артериальной.
Как и у других позвоночных, у круглоротых и рыб имеются так называемые дополнительные сердца, поддерживающие давление в сосудах. Так, в спинной аорте радужной форели есть эластичная связка, выполняющая роль нагнетающего насоса, который автоматически увеличивает циркуляцию крови во время плавания, особенно в мускулатуре тела. Интенсивность работы дополнительного сердца зависит от частоты движений хвостового плавника.
У двоякодышащих рыб появляется неполная перегородка предсердия. Это сопровождается и возникновением легочного круга кровообращения, проходящего через плавательный пузырь, превращенный в легкое.
Сердце рыб гораздо меньше и слабее, чем сердце наземных позвоночных. Масса его обычно не превышает 2,5%, в среднем 1% массы тела, тогда как у млекопитающих оно достигает 4,6%, а у птиц даже 16%.
Кровяное давление (Па) у рыб низкое—2133,1 (скат), 11198,8 (щука), 15998,4 (лосось), тогда как в сонной артерии лошади — 20664,6.
Невелика и частота сокращений сердца—18 .30 ударов в минуту, причем она сильно зависит от температуры: при низкой температуре у рыб, зимующих на ямах, она уменьшается до 1 .2; у рыб, переносящих вмерзание в лед, пульсация сердца на этот период прекращается.
Количество крови у рыб меньше, чем у всех остальных позвоночных животных (1,1, 7,3% массы тела, в том числе у карпа 2,0 .4,7%, сома—до 5, щуки—2, кеты--1,6, тогда как у млекопитающих—6,8% в среднем). Это связано с горизонтальным положением тела (нет необходимости проталкивать кровь вверх) и меньшими энергетическими тратами в связи с жизнью в водной среде. Вода является гипогравитационной средой, т. е. сила земного притяжения здесь почти не сказывается.
Морфологическая и биохимическая характеристика крови различна у разных видов в связи с систематическим положением, особенностями среды обитания и образа жизни. Внутри одного вида эти показатели колеблются в зависимости от сезона года, условий содержания, возраста, пола, состояния особей. Эритроциты рыб крупнее, а их количество в крови меньше, чем у высших позвоночных, лейкоцитов же, как правило, больше. Это связано, с одной стороны, с пониженным обменом рыб, а с другой—с необходимостью усилить защитные функции крови, так как окружающая среда изобилует болезнетворными организмами. В 1 мм3 крови количество эритроцитов составляет (млн): у приматов—9,27; копытных—11,36; китообразных— 5,43; птиц—1,61 .3,02; костистых рыб—1,71 (пресноводные), 2,26 (морские), 1,49 (проходные).
Количество эритроцитов у рыб колеблется в широких пределах, прежде всего в зависимости от их подвижности: у карпа— 0,84 .1,89 млн/мм3 крови, щуки—2,08, пеламиды—4,12млн/мм3. Количество лейкоцитов составляет у карпа 20 .80, у ерша — 178 тыс/мм3. Лейкоциты рыб отличаются большим разнообразием. У большинства видов в крови имеются и зернистые (нейтрофилы, эозинофилы), и незернистые (лимфоциты, моноциты) формы лейкоцитов. Преобладают лимфоциты, на долю которых приходится 80 .95%, моноциты составляют 0,5 .11%, нейтрофилы—13 .31%. Эозинофилы встречаются редко. Например, они есть у карповых, амурских растительноядных и некоторых окуневых рыб.