Страница
3
Открытие в 1953 г. структуры и механизма функционирования ДНК в качестве носителя наследственной информации является началом современного этапа в изучении нуклеиновых кислот. Расшифровка строения нуклеиновых кислот, понимание их функции способствовали значительному прогрессу в изучении белкового синтеза.
В 50—х. годах ХХ в. в опытах с модельными системами было доказано, что местом синтеза белка в клетке являются рибосомы. В это же время была открыта транспортная РНК и установлена вся последовательность этапов биосинтеза белковых молекул.
В середине 50—х. годов на основании исследованний, проведенных в различных разделах биологии, была сформулирована концепция о том, что гены являются участками молекулы ДНК, в которых наследственная информация закодирована чередованием пар нуклеотидов. Был создан новый раздел генетики - молекулярная генетика.
В середине 60-х годов исследователи Р. Холли, Г. Цахау, А. А. Баев и другие разработали принцип определения последовательности нуклеотидов в РНК, который лег в основу изучения структурно-функциональной организации отдельных РНК.
В 1961 г. Ф. Крик сформулировал основные свойства генетического кода. Он математически и экспериментально доказал существование кодирования генетической информации.
В 1961 г. Ф. Жакоб и Д. Моно установили общий принцип работы оперона — группы генов, определяющих синтез функционально связанных ферментов. Эта модель явилась мощным стимулом в разработке практического использования знаний о нуклеиновых кислотах, включая и развитие генной инженерии.
В 1965 г. Ф. Крик выдвинул гипотезу неоднозначного соответствия. Развитие этой гипотезы вылилось в разработку общих принципов структуры генетического кода, что позволило объяснить механизм соответствия кодонов аминокислотам. Одновременно М. Ниренберг и его сотрудники экспериментально доказали, что генетический код содержит равнозначные, как бы взаимозаменяемые кодоны. Они обнаружили, что фенилаланиновая транспортная РНК может присоединяться к двум различным кодонам: как к кодону урацил – урацил - урацил, так и к кодону урацил – урацил – цитозин.
В 1956 г. Д. Тжио и А. Леван установили, что хромосомный набор человека состоит из 46 хромосом. Эта дата считается датой рождения современной цитогенетики человека. К тому времени уже успешно культивировались клетки вне организма, применялись особые способы получения пригодных для проведения хромосомного анализа метафазных пластинок, т. е. пластов клеток, находящихся в метафазе.
В это же время ученые начали обстоятельно исследовать хромосомные аномалии с помощью новых методов, основанных на последних достижениях в изучении структуры нуклеиновых кислот. Было установлено, что хромосомные болезни начинают проявляться уже на самой ранней стадии развития зародыша, а некоторые хромосомные нарушения вызывают его гибель. Накопление знаний о нуклеиновых кислотах и последствиях изменений их структуры в 60—70-х годах привело к появлению в разных странах медико-генетических учреждений для оказания медицинской помощи лицам, страдающим наследственными заболеваниями.
Начиная с 70-х годов ХХ в. разработка методов дифференциальной окраски хромосом для клинической цитогенетики позволила выявить каждую хромосому в ядре клетки, дала возможность во многих случаях судить о происхождении аномальных хромосом. Успех клинической цитогенетики продолжает развиваться и в настоящее время: выделены редкие наследственные заболевания, обусловленные нарушением отдельных сегментов хромосом, а число известных наследственных признаков у человека к 1978 г. достигло 2811.
Таким образом, история развития знаний о нуклеиновых кислотах наглядно демонстрирует постепенный переход от феноменологии нуклеиновых кислот к их экспериментальному и теоретическому изучению с последующим выходом в практику.
Глава 2.Дезоксирибонуклеиновые кислоты.
СОСТАВ И СТРУКТУРА ДЕЗОКСИРИБОНУКЛЕИНОВОЙ КИСЛОТЫ.
Молекулы ДНК, выделенные из ядер клеток, в электронном микроскопе представлены в виде длинных нитей, состоящих из большого числа дезоксирибонуклеотидов. Нити ДНК толще и длиннее, чем нити белков. Длина молекулы ДНК достигает сотен тысяч нанометров. Это несравнимо больше самой крупной белковой молекулы, которая в развернутом виде достигает в длину не более 1ОО—200 нм. Молекула ДНК по массе достигает 6*1О-12 грамма.
Генетическая информация, заключенная в ДНК, состоит из последовательности нуклеотидов. ДНК состоит в основном из четырех нуклеотидов, которые соответствуют четырем азотистым основаниям: аденину, гуанину, тимину и цитозину. Кроме этих оснований, препараты ДНК могут содержать метилированные производные этих оснований.
Основную структурную цепь молекулы ДНК образуют последовательно соединенные друг с другом молекулы пентозы и ортофосфорной кислоты. Цепь ДНК представляет углеводно-фосфатную последовательность, с которой соединены азотистые основания. Углеводные и фосфатные группы выполняют только структурную функцию. Молекулы ортофосфорной кислоты соединяют между собой молекулы дезоксирибозы за счет образования химических связей. При взаимодействии гидроксильной группы 3-го атома углерода одной молекулы пентозы с гидроксильной группой 5-го углеродного атома другой молекулы пентозы отщепляется молекула воды. Тогда у остатков ортофосфорной кислоты сохраняется еще по одной гидроксильной группе, способной диссоциировать. Это обусловливает кислотные свойства всей макромолекулы ДНК.
Молекула ДНК имеет две цепи нуклеотидов, расположенных параллельно друг другу, но в обратной последовательности. Эти цепи удерживаются между собой за счет водородных связей между парами аденин - тимин и гуанин - цитозин. При этом азотистые основания располагаются внутри спирали. Водородные связи образуются между любым электроотрицательным атомом, например кислородом тимина или азотом аденина и атомом водорода, ковалентно связанным с другим электроотрицательным атомом:
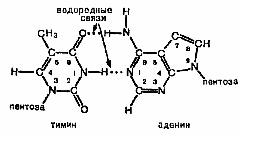
Между аденином и тимином образуются две водородные связи, между гуанином и цитозином — три. Эти связи играют очень важную роль в поддержании вторичной структуры ДНК.
Дополнение аденина тимином и гуанина цитозином, иначе называемое комплементарностью, обеспечивает одинаковое по всей длине двойной спирали расстояние между цепями и образование между противоположными основаниями максимального числа водородных связей, что придает молекуле одновременно устойчивость и подвижность. Последовательность оснований в одной цепи ДНК строго соответствует последовательности оснований в другой цепи. Это является необходимым условием функционирования ДНК и передачи наследственной информации. При необходимости двойная спираль ДНК легко рвется под действием фермента дезоксирибонуклеазы.
Молекула ДНК в ядре клетки не существует изолированно сама по себе. Она окружена связанными с ней белками. Но белки не принимают участия в передаче наследственной информации.
Основными белками, локализованными в ядре клеток и связанными с ДНК, являются специальные белки, называемые гистонами. Гистоны обладают основными (щелочными) свойствами благодаря высокому содержанию в них основных аминокислот. По-видимому, их действие компенсирует в некоторой степени кислотные свойства нуклеиновых кислот. По преобладающему содержанию аминокислот выделяют пять важнейших гистонов: гистон Н1 имеет высокое содержание лизина, гистон Н2b лизина содержит меньше, чем предшествующий гистон, гистон Н2а имеет высокое содержание лизина и аргинина, гистон Н3 содержит большое количество аргинина, гистон Н4 богат аргинином и глицином.