Старая гипотеза «перевернутости» хордовых подтверждается
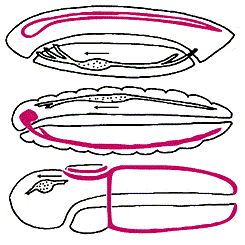
Сопоставление схем организации животных разных типов (сверху вниз):
хордовых (ланцетника), членистоногих и полухордовых.
Передний конец тела всюду обращен влево, спинная сторона - кверху.
Центральная нервная система выделена цветом,
направление кровотока показано стрелкой рядом с сердцем.
Жоффруа Сент-Илер отмечал обратное, по сравнению с беспозвоночными, расположение нервной системы и сердца у хордовых. Но расположение нервной системы - критерий недостаточно убедительный: у беспозвоночных может быть несколько продольных нервных стволов, причем не только на брюшной стороне, но также по бокам и вдоль спины. Например, у представителей класса кишечнодышащих из типа полухордовых (ближайших родственников хордовых) два продольных нервных ствола: брюшной и спинной, причем спинной ствол у них частично свернут в трубку, точно так же, как у хордовых. Следовательно, можно предположить, что у предка хордовых было два нервных ствола, как у современных полухордовых, но затем брюшной ствол редуцировался и остался только спинной. И в этом случае гипотеза “перевернутости” не нужна.
Расположение сердца - признак более надежный. Заметим, что направление кровотока у кольчатых червей, членистоногих, полухордовых и хордовых (если последних перевернуть) совпадает. Однако многие беспозвоночные лишены кровеносной системы, поэтому считается, что разное расположение сердца (снизу у хордовых и сверху у беспозвоночных) связано с независимым возникновением у них кровеносной системы.
Среди других анатомических признаков, подтверждающих “перевернутость” хордовых, отметим лишь расположение печени: над средней кишкой у беспозвоночных и под - у хордовых [6].

Расположение желтка (показано цветом) у зародышей лягушки (вверху) и дрозофилы.
Передний конец тела обращен влево, спинная сторона - вверх.
Кроме того, гипотеза “перевернутости” хордовых подтверждается в результате сравнения богатых желтком зародышей наземных членистоногих и таких позвоночных, как рыбы, земноводные, рептилии и птицы. У членистоногих желток всегда располагается на спинной стороне зародыша, а у позвоночных - всегда на брюшной [7]. Строение таких зародышей становится сопоставимым, только если признать гомологию спинной стороны позвоночных брюшной стороне беспозвоночных.
Молекулярная биология развития
В последние годы убедительные доказательства гипотезы “перевернутости” хордовых получены молекулярными биологами [8]. У зародышей позвоночных (лягушки и рыбы) на стадии гаструлы на спинной стороне синтезируется белок CHD (chordin), а на брюшной - белок BMP-4 (bone morphogenetic protein-4). У насекомых (дрозофилы) на стадии гаструлы на спинной стороне синтезируется белок DPP (decapentaplegic), а на брюшной - белок SOG (short gastrulation).
Формирование структур и органов спинной и брюшной сторон тела у позвоночных и насекомых определяется соотношением концентраций упомянутых белков. Так, у зародыша лягушки в норме белок CHD синтезируется на спинной стороне, но если в опыте повысить его концентрацию на брюшной с помощью инъекции, необходимой для его синтеза матричной РНК, то на этом месте в дальнейшем разовьются структуры спинной стороны. Наоборот, искусственное повышение концентрации белка BMP-4 на спинной стороне зародыша вызывает развитие там структур брюшной стороны. Аналогичные результаты были получены и на дрозофиле: формирование структур спинной стороны связано с повышенной концентрацией белка DPP, а брюшной - SOG.
Выяснилось также, что в белках CHD и SOG оказались одинаковыми 28% аминокислотных последовательностей, а из 100 аминокислот C-концевых частей молекул белков BMP-4 и DPP, отвечающих за их функционирование, совпадают 76%. Даже 28%-ая гомология сравниваемых белков свидетельствует о несомненном общем происхождении, от одного и того же белкового предка. Таким образом, “спинной” CHD позвоночных гомологичен “брюшному” SOG дрозофилы, а “брюшной” BMP-4 позвоночных гомологичен “спинному” DPP дрозофилы. Но ведь эти гомологии - не что иное, как подтверждение гипотезы “перевернутости” хордовых!
Удивительна эволюционная консервативность функции рассматриваемых белков, которая проявляется в том, что белки лягушки CHD и BMP-4 функционально активны в зародыше дрозофилы, а белки дрозофилы SOG и DPP - в зародыше лягушки. Экспериментально показано, что “брюшной” белок лягушки BMP-4 вызывает у зародыша дрозофилы формирование структур спинной стороны, а “спинной” белок лягушки CHD приводит к формированию зародышем дрозофилы структур брюшной стороны. Наоборот, белки дрозофилы - “брюшной” SOG и “спинной” DPP - соответственно вызывают формирование зародышем лягушки структур спинной и брюшной сторон. А ведь, согласно современным оценкам, с момента расхождения эволюционных линий, ведущих к насекомым и хордовым, прошло более полумиллиарда лет! Такая консервативность белков делает гипотезу “перевернутости” хордовых еще более правдоподобной.
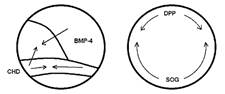
Диффузия (направление показано стрелками) белков, определяющих в начале гаструляции формирование спинной и брюшной сторон у зародышей позвоночных и насекомых: лягушки (слева) и дрозофилы. У зародыша лягушки спинная сторона определяется повышенной концентрацией белка CHD в области так называемого организатора (спинной губы бластопора), а брюшная - BMP-4 на противоположной организатору стороне зародыша. У зародыша дрозофилы спинная сторона определяется повышенной концентрацией белка DPP и брюшная - SOG. Так как в парах CHD/SOG и BMP-4/DPP белки гомологичны, можно сделать вывод, что спинная сторона лягушки соответствует брюшной стороне дрозофилы и, наоборот, брюшная сторона лягушки соответствует спинной стороне дрозофилы. Эти гомологии подтверждают гипотезу “перевернутости” хордовых.
Существуют и другие, дополнительные свидетельства в пользу гипотезы, полученные в результате сопоставления некоторых генов, из которых мы упомянем лишь о двух [9]. Развитие продольного нервного ствола связано с активностью вдоль его средней линии гомологичного для разных животных (насекомых, позвоночных, круглых червей) гена netrin: соответственно положению нервной системы этот ген активен на брюшной стороне у насекомых и на спинной - у позвоночных. Таким образом, брюшной нервный ствол беспозвоночных гомологичен спинному стволу позвоночных. Развитие сердца у дрозофилы определяется синтезом белков TINMAN и DMEF2, а у позвоночных - гомологичных им белков. Следовательно, расположенное на спинной стороне сердце насекомых и на брюшной стороне сердце позвоночных также оказываются гомологичными.
Итак, благодаря современным данным молекулярной биологии, сравнительной анатомии и эмбриологии подтвердилась гипотеза “перевернутости” хордовых, выдвинутая Жоффруа Сент-Илером еще в начале прошлого века.
Список литературы